


Полная версия
Парадоксы эволюции. Как наличие ресурсов и отсутствие внешних угроз приводит к самоуничтожению вида и что мы можем с этим сделать




Рис. 4. Повторное заполнение во влажную фазу: граница раздела фаз проходит по новой линии, в результате чего часть ячеек с щелочными отложениями на стенках заполняется кислой средой и наоборот

Рис. 5. После нескольких циклов смены сухих и влажных фаз часть ячеек оказывается заполненными разнородными слоями, с возможным формированием биоэлектрического «вольтового столба» – аккумулятора энергии, а сами ячейки окружены ячейками с содержимым существенно иного рН

Рис. 6. Мембраны «научились» энергетически самоподдерживаться и самовосстанавливаться, возможно, с помощью структурных нуклеиновых кислот и рибозимов. Минеральная составляющая полупроницаемых мембран может впоследствии «вымываться», оставляя ассоциированные с новыми органическими оболочками каталитические железосерные кластеры
На близком принципе – чередования соленых и пресных фаз в изолированных элементах – уже построен ряд экспериментальных энергетических установок. С их помощью предполагается использовать «энтропийную» энергию градиента солености Мирового океана, оцениваемую до 1700 ТВт*ч/год (Skilhagen S.E., 2011), так называемую «голубую энергию» океана. Интересными прототипами можно назвать емкостную технологию итальянского инженера-физика Дориано Броджиоли (Doriano Borgioli, 2009) на основе двуслойного электролитического конденсатора (ионистора) большой емкости, работающего по сложному циклу заряда/разряда и поочередного наполнения камеры конденсатора соленой и пресной водой, и более продвинутую технологию группы исследователей из Стэнфордского университета (Ye M. et al., 2019), использующую «батареи энтропийного смешивания» (БЭС; Mixing Entropy Batteries, MEB). БЭС включает два электрода с большой поверхностью контакта, между которыми происходит чередование фаз морской и пресной воды. Один электрод состоит из берлинской лазури, смеси нескольких гексацианоферратов (II), весьма простых соединений железа, азота и углерода, другой представляет собой проводящий органический полимер, полипиррол. В циклическом процессе при заполнении камеры батареи пресной водой ионы натрия и хлориды выходят из соответствующих электродов в воду, создавая электрический ток между электродами. При смене пресной воды на морскую ионы натрия и хлориды забираются обратно в электроды, также образуя электрический ток, но уже обратной направленности.
Технически протонный градиент между средами с разным рН может быть заменен на натриевый и/или калиевый градиенты, имеющие бОльшую буферную емкость из-за большего содержания калия и натрия в рассматриваемых водах, нежели свободные протоны. На идее первичного калиевого градиента, наследуемого почти всеми формами жизни – преобладания калия внутри, а натрия снаружи клетки – основывается модель Армена Мулкиджаняна, Дарьи Дибровой, Михаила Гальперина и Евгения Кунина (Mulkijanian A. et al., 2012; Диброва Д. и соавт., 2015). В этой модели за основу для формирования протобиологических структур принимается конденсат испарений наземных геотермальных полей, в котором, как и в клеточных цитоплазмах, преобладает калий. Сам пресный конденсат неизбежно должен был находиться в щелочном окружении богатых натрием горячих геотермальных вод. При наличии адекватного полупроницаемого разделения на этой основе могла возникнуть первичная фосфатная, а затем самоподдерживающаяся натрий-калиевая мембранная энергетика. Как показывают недавние биоинформационные исследования группы Мулкиджаняна (Козлова М. И. и соавт., 2020), эта древнейшая натриевая энергетика («натриевый мир») не заместилась полностью более «современной» протонной энергетикой, а оказалась органично встроенной в конфигурацию биоэнергетических и биоинформационных процессов большинства современных эукариот, архей и бактерий. Даже у высших животных, включая человека, ключевые белки межклеточных коммуникаций – ассоциированные с G-белком рецепторы (GPCR, G-protein coupled receptors), одна из самых широко представленных в организме групп белков, – имеют прямое происхождение от древнейших белковых энергетических структур «натриевого мира».
Строительные элементы доклеточных структур
Было бы любопытно предположить, какие органические полимеры могли составить первичную органическую основу многослойных отложений на минеральных мембранах, обеспечив функциональный базис дальнейшего развития. Липиды и полисахариды, составляющие такую основу для большинства современных мембран живых организмов, возможны, но их синтез требует достаточно изощренных каталитических механизмов, практически невероятных на данном этапе развития биологической сложности. Белки также вполне могут подойти на эту роль, так как принципиально способны образовывать стабильные оболочки живых структур и способны к автокаталитическому самоподдержанию. Потенциальным недостатком может оказаться тот факт, что белки с (почти) случайной последовательностью аминокислот (в отсутствие еще носителей наследственной информации) чаще всего будут оказываться структурно неустойчивыми и неспособными к самоукладке в четвертичные (многомолекулярные) макрокомплексы. Для нуклеиновых кислот структурная функция не рассматривается в качестве первичной даже на ранних этапах эволюции. Обычно предполагается, что нуклеиновые кислоты эволюционно начинают как носители наследственной информации и катализаторы первых биохимических реакций. Однако и сейчас, в современных организмах, нуклеиновые кислоты зачастую выполняют структурную функцию даже у эволюционно далеких организмов, причем, и очень часто, в экстремальных для них условиях. При этом выполнение такой структурной функции нуклеиновыми кислотами не демонстрирует критической зависимости от четкой нуклеотидной последовательности. Так, нуклеиновые кислоты составляют основу гнойного содержимого ран – практически последней линии защиты при атаке патогенами физических повреждений многоклеточных организмов. Видимо, совсем неслучайно именно это содержимое стало местом открытия самих нуклеиновых кислот (ДНК) швейцарским врачом Иоганном Фридрихом Мишером в 1869 году.
Физиологической основой формирования воспалительных структур, основанных на ДНК – так называемых внеклеточных нитей нейтрофилов (NET, neutrophil extracellular traps), является процесс нетоза нейтрофилов. Нейтрофилы – наиболее распространенная разновидность лейкоцитов, рядовые солдаты острой воспалительной реакции организма. Сам нетоз можно считать разновидностью апоптоза – запрограммированной клеточной смерти, а нейтрофилы – клетками-камикадзе, предназначенными умереть, отдав самое святое – свою ДНК – в качестве кирпичей на строительство оборонительных рубежей всего организма (собственно смерть нейтрофила при этом не является совсем обязательной, а сам нетоз может создавать при определенных условиях и существенные проблемы для всего организма (Papayannopoulos V., 2018)). Важность этого защитного механизма, имеющего, вероятно, очень глубокие эволюционные корни – своего рода возврат к первичным основаниям жизни, своеобразное повторение филогенеза в патогенезе, подтверждается тем фактом, что наличие ферментов экзонуклеаз, способных разрушать NET структуры, служит существенным фактором вирулентности бактерий (Sharma P. et al., 2019). Напротив, и сами бактерии способны формировать из ДНК несущую решетку своего внеклеточного матрикса – основу бактериальных биопленок, своеобразный защитный чехол бактериального сообщества. Причем ключевые единицы такой ДНК-решетки аналогичны так называемым структурам Холидея – крестообразным формированиям двухцепочечной ДНК (рис. 7). И совершенно неслучайно именно ДНК-структуры, в том числе сформированные с использованием структур Холидея, применяются в качестве строительных блоков для ряда нанотехнологических материалов. Тем более что ДНК в качестве структурного материала обладает рядом уникальных свойств: например, высокоточной самосборкой и самоукладкой в полимолекулярные комплексы (Rothemund P. W. K., 2006), способностью проводить ток (в том числе фотохимического происхождения), что может быть весьма существенным фактором для участия в энергопроизводящих ячеистых структурах (см. рис. 2-6). Причем менее организованная, но более компактная А-форма (конформационно близкая к двухцепочечной РНК и ДНК-РНК гибридам) делает это на порядок более эффективно, чем более известная В-форма – правозакрученная спираль (Artes J. M. et al., 2015).
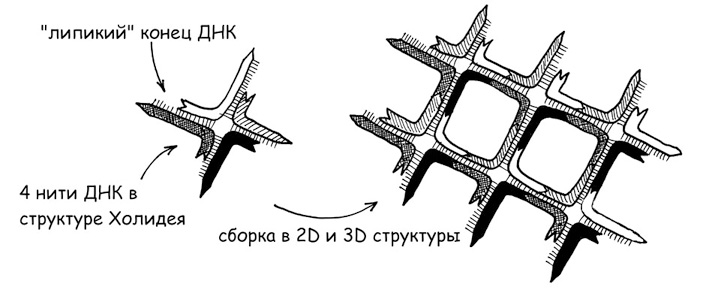
Рис. 7. Структуры Холидея и их искусственные производные
Рассматривается, что именно ДНК-РНК гибриды и могли быть первыми протобиологическими молекулами: Цзяньфен Сю и Джон Сазерленд с коллегами (Jianfeng Xu, John Sutherland et al., 2020) показали, что в кислой среде, в присутствии ионов натрия, магния, хлоридов и нитритов из простейших неорганических веществ типа цианидов может образовываться система нуклеозидов из 4 оснований, 2 из которых – пуриновые дезоксирибонуклеозиды (аденозин и инозин), и 2 пиримидиновые рибонуклеозиды (цитидин и уридин). До этого не удавалось продемонстрировать возможность возникновения рибозы и рибозидов в условиях первичной Земли (в отличие от дезоксирибозы и ее производных). В последнее время, при почти полном доминировании идеи РНК-мира (первичной роли РНК в возникновении жизни, ввиду ее способности как к сохранению информации, так и к катализу, в том числе способствующему саморепликации), появляются и другие свидетельства того, что РНК и ДНК, а возможно, и их гибриды, появились и могли сосуществовать еще до возникновения жизни (Extance A., 2020). В качестве одного из таких вариантов японскими исследователями из университета Нагоя (Murayama K. et al., 2021) предложены близкие к ДНК достаточно стабильные ксенонуклеиновые кислоты (КНК), в частности на основе L-треонинола (L-aТНК) – алифатической нуклеиновой кислоты (то есть без пуринового или пиримидинового колец, свойственных «нормальным» нуклеиновым кислотам). Особенностью полимеров данной кислоты, возможно, даже с включением «нормальных» нуклеотидов, является возможность самосборки (автополимеризации) в виде двойных цепей без участия белковых ферментов. Более того, фрагменты КНК способны далее сами выступать в качестве катализаторов самосборки, подобно некоторым РНК-фрагментам (рибозимам). Дополнительным свидетельством возможности сценария первичных структурных нуклеиновых кислот может служить обнаружение конъюгатов углеводов с некодирующими РНК – гликан-РНК – в составе мембран ряда организмов, где они, похоже, выполняют роль сигнальных рецепторов (Flynn R. A. et al., 2021). Этим самым открыт своеобразный мостик, возможно, очень древний, между миром генетической информации нуклеиновых кислот и миром разделительных и сигнальных углеводных структур.
Такие структуры возможны?
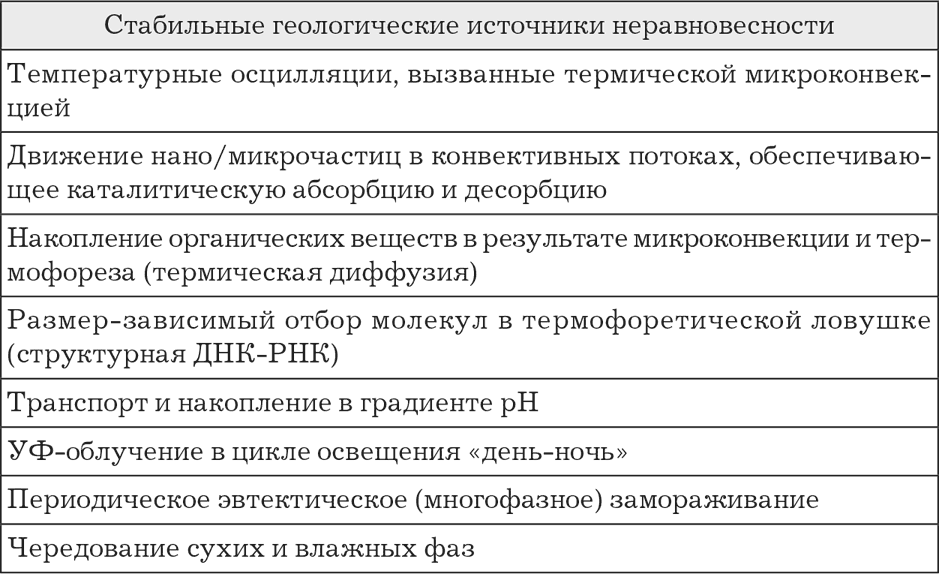
Важность разделительных структур в эволюции нуклеиновых кислот акцентируется в работах немецкого исследователя Кристофа Маста (Christof Mast et al., 2010; 2013). Мастом сформулирована теория термодинамической ловушки. Она показывает, что наличие термохимических градиентов создает дополнительные состояния неравновесности, стимулирующие в частных случаях полимеризацию, репликацию и концентрирование ДНК и возникновение дарвинского отбора в более общих случаях (табл. 1). Помимо термического градиента, вызванного геологическими условиями (разница температур гидротермального источника и внешней (например, морской воды), дополнительный, хотя и сравнительно микроскопический, но в некоторых ситуациях решающий вклад мог вносить и химический градиент протонов (электронов) и/или натрия, способный, в принципе, также создавать температурный градиент. Этот искусственно возникающий градиент, в свою очередь, несмотря на свою кажущуюся незначительность, мог способствовать впоследствии независимости протобиологических структур от геологических. По некоторым расчетам, даже незначительного температурного градиента в отдельных случаях может оказаться достаточно, например для обеспечения простого деления протоклеток: перемещение более «горячих» молекул двуслойной наружной мембраны наружу, движимое простыми физическими механизмами, увеличивает среднюю кривизну и усиливает любое локальное сжатие протоклетки вплоть до полного разделения на две (Romain Attal and Laurent Schwartz, 2021).
С другой стороны, мы видим, как уже на самых ранних этапах возникает двухвариантность реализации химического (протонного) градиента: 1) как концентрирование в форме химических связей (или в составе любых синтезируемых соединений, или, что оказывается выгоднее, в форме нескольких универсальных «энергетических валют», например АТФ); 2) как рассеивание (диссипация) энергии для более общей неспецифической модификации условий окружающей среды («контекста организации»).
Таблица 1. Примеры состояний геологической неравновесности (по Mast C. et al., 2010)
Насыщенные нуклеиновыми кислотами неорганические мембраны могли участвовать в трансмембранном и трансклеточном переносе энергии, что запустило бы эволюционный отбор нуклеиновых кислот. Впоследствии же возникший отбор нуклеиновых кислот с первоначально случайными последовательностями смог бы выявить ряд комбинаций, оказавшихся способных к саморепликации, и сделал «чистые» ДНК и РНК ключевыми операторами наследственной информации со своим четко определенным функционалом. В этом смысле первыми автореплицирующимися структурами могли быть общие предки вирусов и прокариот, паразитировавшие на длинных структурных/проводящих ДНК. Чередование кислотных и щелочных, влажных и сухих фаз в существовании доклеточных протобиологических структур можно сопоставить с чередованием хаотической и динамической фаз в теории эволюции информационных систем. Учитывая бóльшую чувствительность РНК к рН среды, щелочные влажные фазы могли выполнять роль хаотического «перемешивающего слоя» в протоэволюции нуклеиновых кислот и в большей степени могло быть связаны с РНК, а в кислотные или сухие фазы более стабильные ДНК фиксировали достижения и канализировали развитие (БОН: глава IV). Д. С. Чернавский, физик и математик, автор статистической теории «перемешивающих слоев», исходя из физических и информационных представлений, также считал ДНК более предпочтительной в качестве приоритетной молекулы при возникновении биологической сложности.
Есть основания предполагать, что эволюция протоживого и живого первые миллионы, если не миллиарды лет – от первых, еще не идентифицированных самореплицирующихся молекул до возникновения уцелевших до наших дней, хоть и в довольно измененном виде, архей и бактерий – проходила в значительной степени в привязке к горячим щелочным термальным источникам, подводным или наземным, являвшимися в первую очередь источниками химической энергии и минимальной упорядоченности, негэнтропии, которую и научились «есть» первые организмы.
В зависимости от того, с какой стороны кислотно-щелочного раздела им оказалось сподручнее усваивать эту негэнтропию, то есть в какую сторону оказался направлен протонный (протонно-натриевый?) насос – на выталкивание или запуск протонов в клетку – первые организмы разделились на линию бактерий (выталкивание протонов) и линию архей (запуск в клетку). Представляется, что самым ранним протобактериям и протоархеям энергетически выгоднее было бы сосуществование; само отнесение к протоархеям и протобактериям могло оказываться достаточно условным: направленность транспорта протонов в конкретной ячейке/протоклетке могла меняться в зависимости от изменения условий окружения в циклической геологической системе. Совершенствование биохимических механизмов в любом случае позволило им в конце концов разделиться и стать полностью самостоятельными. Уже самые древние независимые археи и бактерии имели между собой значительные различия как в ключевых наследственных генетических механизмах, так и структуре – в первую очередь в строении наружных оболочек на основе липидных полупроницаемых мембран. И когда этим двум сильно разошедшимся формам жизни вновь оказалось выгодным заново начать жить вместе в странном симбиозе, то, несмотря на громадные различия, накопившиеся за почти два миллиарда лет раздельного существования, позволившие успешно произойти этому событию только один раз и предопределившие все последующие проблемы совместного существования базовые энергетические механизмы археи и бактерии в новом симбиозе снова подошли друг к другу как ключ к замку. Бактерии, казалось бы, заняли в этом симбиозе подчиненное, «крепостное» положение маленьких органелл в большой архейной протоэукариотической клетке – с вроде бы как ограниченной ролью маленьких «электростанций», с невероятной эффективностью вырабатывающих огромное количество необходимой клетке энергии. Это сожительство дало начало как большей части наблюдаемой эволюции сложной жизни, так и ее предопределенной смерти.
Жизнь бактерий в архейном замке
Работа их наследников – митохондрий – в сущности сохранила принципиальную схему выработки энергии, предполагаемую для первичных форм жизни в морских подводных щелочных гидротермальных источниках:
1. Наличие источника протонов (ионов водорода) и/или ионов натрия – для первичных форм жизни это окружающая среда: богатая протонами (то есть кислая) и соленая (то есть богатая натрием) морская вода или соленая вода закрытых озер; в митохондриях это узкое пространство между наружной и внутренней мембранами митохондрий. Сюда изнутри митохондрий выталкивает протоны цепь белковых комплексов, расположенных на внутренней мембране. Выталкивание происходит за счет движения электронов по этой цепи – так называемой дыхательной или электрон-транспортной цепи (ЭТЦ). Электроны и протоны берутся из происходящего внутри митохондрий цикла ферментативных реакций (цикла Кребса), полностью, до углекислого газа и воды, разлагающего пируват – продукт распада глюкозы в гликолизе. Электроны по ЭТЦ движутся к своему «приемному пункту» – конечному акцептору электронов, окислителю. Реакции приема-передачи электронов соответственно называются окислительно-восстановительными. У животных конечным приемщиком (акцептором) электронов служит кислород, но есть одноклеточные (например, некоторые бактерии), у которых электроны принимаются серой, нитратами, железом и другими веществами.
2. Наличие бедной протонами (то есть щелочной) среды – для первичных форм жизни это щелочная вода источников, в митохондриях это внутренний матрикс с ферментами цикла Кребса, откуда по ЭТЦ наружу выкачиваются протоны.
3. Наличие полупроницаемой мембраны между указанными выше средами. Полупроницаемая – это значит, что для прохождения через нее вещества должны потратить энергию или совершить работу (или, как обычно бывает в жизни, кто-то должен сделать это за них). В данном случае работа может совершаться за счет разницы концентраций протонов или натрия по разные стороны мембраны (что и образует уже названные протонный или натриевый потенциалы). Для первичных форм жизни мембраны могли быть неорганические, например серпентинитовые, с каталитическими железосерными кластерами, для современных форм жизни характерны липидные, в которые встроены многочисленные каталитические белки – ферменты, включая белки ЭТЦ. По сути, как говорилось, сам по себе потенциал уже является формой энергии. В развитых формах жизни этот потенциал «заряжает» универсальный биологический аккумулятор, то есть благодаря ему на окончании цепочки ЭТЦ в процессе упомянутого окислительного фосфорилирования образуется АТФ из АДФ. Для первичных форм жизни вопрос транспорта электронов учеными еще не решен: его постоянный направленный механизм обязан был существовать, чтобы обеспечить образование первичных, самых простых органических молекул, например из углекислого газа, самого распространенного во все времена источника углерода. Но что стало самым первым источником электронов – восстановителем – в цепи реакций, в результате которой могли бы образовываться хотя бы самые простые органические молекулы: метан, формальдегид, цианид и им подобные простые вещества? Теоретически им мог стать тот же молекулярный водород Н2, но практически пока не удается продемонстрировать реальность подобной реакции. Известный ученый и популяризатор науки Ник Лейн (2015) очень упорно, но, как многим представляется, недостаточно убедительно, настаивает, что подходящие условия для вовлечения водорода в эту реакцию могли возникнуть именно в щелочных гидротермальных источниках с участием железосерных кластеров. Других приличных вариантов первичного запуска транспорта электронов в любом случае пока еще нет. Для нас же важна уверенность многих ученых, что в любом случае эволюция живого от первых до-жизненных форм до сложившихся клеток происходила в щелочных термальных источниках или в непосредственной связи с ними, сохраняя на этом отрезке развития общие принципы получения, удержания и использования энергии.
Когда приблизительно 1,5–2 миллиарда лет назад бактериальные предки митохондрий начали свой переезд в живой «замок» древней археи, они, разумеется, прихватили вместе с собой весь свой небогатый скарб: не ахти какой, но все-таки своеобразный биологический «капитал» – свой наследственный материал.
Но со временем необходимость поддержания стабильности «главного» генома археи (а по факту – уже общей наследственности) предопределила перемещение и почти всего митохондриального генома во вновь возникшее ядро – ризницу хранения священного наследственного материала в главном соборе усложнившегося архейного-эукариотического замка (БОН: глава VII). Однако проблемы качественного управления на месте (гемба-менеджмента) заставили несколько последних генов (у человека – 13 генов белков, 22 транспортных РНК и 2 рибосомальных РНК) до конца оставаться непосредственно в самих митохондриях. В конечном счете та же необходимость уменьшения митохондриальной гетероплазмии, то есть спонтанно возникающего в жестких биохимических условиях внутриклеточной водородной «электростанции» чрезмерного разнообразия генов – «локальных производственных процедур», совершенно логическим образом вызвало возникновение двух полов с наследованием родительского ядерного материала в пропорции 50 на 50 %, и со 100 % наследованием митохондриального генома от одного материнского родителя. Практически любой сбой в согласованной работе между митохондриальными генами, локализованными в ядре, и локализованными в митохондрионе ведет к падению эффективности работы электрон-транспортной цепи (ЭТЦ), что проявляется увеличением образования активных форм кислорода (АФК), и, соответственно, итоговым уменьшением образования АТФ. Если рассогласованность имеет постоянный характер, это, согласно модифицированной свободнорадикальной теории старения, увеличивает скорость старения и сокращает продолжительность жизни особи (Lane N., 2015).
Опираясь на представление о стержневой роли митохондрий в энергетическом обеспечении как эволюции сложных организмов, так и развития – от рождения до смерти – отдельного организма, можно увидеть глубочайшую вовлеченность митохондрий во все физиологические процессы организма. В своей тяжелейшей и наиболее очевидной форме для человека это проявляется в виде так называемых митохондриальных заболеваний, но участие митохондриона неизбежно просвечивается и во множестве других, самых неожиданных феноменах других патологий.
Когда они ломаются
Собственно, митохондриальными заболеваниями (МЗ) в узком смысле называют болезни, непосредственно связанные с дефектами дыхательной (электрон-транспортной) цепи (ЭТЦ), то есть процесса окислительного фосфорилирования. Например, синдром Лея (Leigh syndrome) – подострая некротизирующая энцефаломиелопатия вследствие появления в стволе мозга, мозжечке, базальных ганглиях очагов некроза, глиоза, прорастания сосудов, проявляющаяся задержкой или даже регрессией психомоторного развития, мышечной гипотонией, на что впоследствии накладываются самые разнообразные психосоматические нарушения, вплоть до судорожных припадков. Связана с функциональной дисфункцией крупных молекул I или IV комплекса ЭТЦ и/или дисфункцией малой молекулы – цитохрома С. МЗ возникают вследствие наследственных или спонтанных мутаций в митохондриальной ДНК (мтДНК) или в ДНК ядерных генов, контролирующих работу митохондрий. Сейчас известно более 300 таких возможных мутаций, затрагивающих около 100 генов. Остаются тем не менее случаи с явной клиникой митохондриальных заболеваний, но без выявленных генетических нарушений. Как правило, МЗ затрагивают только отдельные типы клеток в определенных органах и тканях, преимущественно (но далеко не всегда) с высокими энергетическими потребностями, такими как нервная и мышечная ткани. Тем не менее узкая поражаемость во многих случаях только определенных органов, как, например, оптическая нейропатия Лебера (Leber Hereditary Optic Atrophy, LHON), при которой повреждаются только ганглиозные клетки сетчатки глаза, все еще требует более точного объяснения. Возможно, более системные нарушения митохондриальной функции останавливают жизнь организма еще на уровне первых делений оплодотворенной яйцеклетки, а для реализации скрытых нарушений, которые приводят к органоспецифичным МЗ, требуются существенные ко-факторы, скорее приобретенные, чем наследуемые.

