


Полная версия
Парадоксы эволюции. Как наличие ресурсов и отсутствие внешних угроз приводит к самоуничтожению вида и что мы можем с этим сделать



Аутофагия – ключ к разгадке?
Важной находкой можно считать более высокий уровень аутофагии у некоторых видов летучих мышей, причем как базовой, так и индуцированной (Laing E. D., 2019). Аутофагия – процесс внутриклеточного удаления органелл и макромолекул, в первую очередь поврежденных. Этот феномен у летучих мышей, очевидно, в значительной степени относится к удалению разрегулированных оксидативным стрессом митохондрий. Аутофагия может выполнять как функцию «очищения» клетки, так и питания в условиях голодания. Исследования в области аутофагии находятся в мейнстриме современной молекулярной биологии, что особенно подтвердилось присуждением Есинори Осуми Нобелевской премии в 2016 году именно за выдающиеся работы в этой области (единственная в прошедшем десятилетии Нобелевская премия в области физиологии и медицины, присужденная одному человеку).
Аутофагия у летучих мышей очень активно стимулируется вирусами, и после своего запуска активно предотвращает развитие вирусной инфекции, в том числе в отношении других вирусов. Антивирусное действие аутофагии у рукокрылых опосредовано через механизмы врожденного иммунитета, в целом нетипичные для других млекопитающих, то есть оказываясь еще одним вариантом интерференции.
Аутофагия может выступать альтернативой основному виду программируемой клеточной смерти – апоптозу, «стандартному» пути устранения вирус-инфицированных и перерожденных опухолевых клеток (в отношении опухолевых клеток аутофагия может даже выступать усилителем их выживаемости): механизм аутофагии еще оставляет клетке шанс на выживание, а запущенный механизм апоптоза – уже нет. Необходимо отметить, что было бы опрометчиво голодание животного считать универсальным инструментом некоего «очищения» всего организма путем аутофагии, которая все-таки сама по себе отчетливо внутриклеточные явление, но, безусловно, способное дать выраженные положительные проявления и на уровне организма.
У большинства летучих мышей заблокированы или заторможены традиционные для других млекопитающих механизмы неспецифической противовирусной внутриклеточной защиты: упоминавшееся уничтожение свободной внутриклеточной ДНК (сигнальный путь STING), формирование ключевых внутриклеточных инструментов воспаления – инфламмасом (сигнальные пути NLRP3 и PYHIN/AIM), выработка одного из основных медиаторов воспаления при вирусной инфекции – интерлейкина-1-бета (Irving A. T. et al., 2021). Так как значительная доля свободной ДНК относится к митохондриальной ДНК (мтДНК) – важнейшему средству внутри- и межклеточной коммуникации, то заторможенный путь STING предоставляет митохондриону больше функциональной свободы.
Многоопытные и хитроумные
В очень большом обобщении можно констатировать, что стратегия иммунных процессов у летучих мышей подобна стратегии восточных единоборств или пути Одиссея: уход от прямого противодействия, направление силы противника в свою пользу, навязывание своих условий взаимодействия (рис. 1).

Рис. 1. Многоопытные, хитроумные, рукокрылые
Несмотря на выраженную эффективность метаболических и иммунных механизмов летучих мышей, их удивительную «вписываемость» в самые разнообразные экологические ниши, нельзя сказать, что они совершенно безупречны и обеспечивают мышам универсальную невосприимчивость к инфекциям. В последние годы целые многотысячные, если не многомиллионные колонии североамериканских летучих мышей буквально выкашивает эпидемия синдрома «белого носа». Счет жертв идет уже на десятки миллионов, ставя под угрозу выживания целые популяции и даже виды рукокрылых. Возбудителем заболевания является холодолюбивый грибок Pseudogymnoascus destructans, предпочитающий укромные места с температурой от +4 до +20оС. Такие же места предпочитают для зимней спячки летучие мыши. Здоровые мыши обычно несколько раз за время сна просыпаются не более чем на час, при этом их температура повышается. Заразившись, летучие мыши часто и надолго выходят из спячки, начинают беспокойно летать. Такие интенсивные пробуждения ведут к быстрой потере накопленных жировых запасов. Затем на крыльях возникают плохо заживающие рубцы, на мордочках высыпает белый грибковый налет. В большинстве случаев летучая мышь погибает, не дождавшись времени нормального весеннего пробуждения. Важно отметить, что грибок Ps. destructans, ставший причиной североамериканской эпидемии у летучих мышей, имеет европейское происхождение, а сами европейские летучие мыши вполне устойчивы к этой инфекции. Поэтому даже такая продвинутая система защиты от инфекций, комбинирующая механизмы, связанные как с повышенной, так и пониженной температурой, может быть взломана. И даже неудивительно, что взломщиками оказались грибки, одни из самых изощренных паразитов, на счету которых сотни и тысячи исчезнувших с лица Земли видов, включая, возможно, самых исполинских ее хозяев – динозавров (БОН: глава IX).
Касательно же самих летучих мышей, давших нам первую подводку о связях здоровья и эволюции, можно заключить, что биологические особенности летучих мышей, а именно способность к полету, то есть экстремальной физической мобилизации, устойчивость к множеству опасных для других млекопитающих вирусов, невероятно высокая продолжительность жизни, умение впадать в ступор и спячку, не одна, а несколько «нормальных» температур тела в зависимости от физиологического статуса – все эти особенности в значительной, если не определяющей степени вызваны невероятным взаимодействием необыкновенно точно отрегулированных систем энергетического метаболизма и иммунитета. Краеугольным камнем этого взаимодействия являются митохондрии, удивительные двигатели эволюции эукариот (организмов на основе клеток, обладающих ядром, включая все многоклеточные организмы) и гаранты их здоровья.
Библиографический список
1. Крускоп С. В. Летучие мыши: Происхождение, места обитания, тайны образа жизни. – М.: Фитон XXI, 2013.
2. Wang L.-F., Walker P. J., Poon L. L. M. Mass extinctions, biodiversity and mitochondrial function: are bats ‘special’as reservoirs for emerging viruses? (2011). Curr. Opin. Virol.; 1: 649–657.
3. Calisher C. H., Childs J. E., Field H. E., Holmes K. V., Schountz T. (2006). Bats: important reservoir hosts of emerging viruses. Clin. Microbiol. Rev.; 19: 531–545.
4. Luis A. D., Hayman D. T. S., O’Shea T. J., Cryan P. M., Gilbert A. T., Pulliam J. R. C., Mills J. N., Timonin M. E., Willis C. K., Cunningham A. A., Fooks A. R., Rupprecht C. E., Wood J. L., Webb C. T. (2013). A comparison of bats and rodents as reservoirs of zoonotic viruses: are bats special? Proc Royal Soc B Biol Sci.; 280: 20122753.
5. Mandl J. N., Schneider C., Schneider D. S., Baker M. L. (2018). Going to Bat(s) for Studies of Disease Tolerance. Front. Immunol. 9: 2112.
6. Zhang G., Cowled C., Shi Z., Huang Z., Bishop-Lilly K. A, Fang X., Wynne J. W., Xiong Z., Baker M. L., Zhao W., Tachedjian M., Zhu Y., Zhou P., Jiang X., Ng J., Yang.L, Wu L., Xiao J., Feng Y., Chen Y., Sun X., Zhang Y., Marsh G.A., Crameri G., Broder C. C., Frey K. G., Wang L. F., Wang J. (2013). Comparative analysis of bat genomes provides insight into the evolution of flight and immunity. Science 339: 456–60.
7. Shen Y. Y., Liang L., Zhu Z. H., Zhou W. P., Irwin D. M., Zhang Y. P. (2010). Adaptive evolution of energy metabolism genes and the origin of flight in bats. ProcNatl Acad Sci USA. 107: 8666–71.
8. Ngo J., Osto C., Villalobos F., Shirihai O. S. (2021). Mitochondrial Heterogeneity in Metabolic Diseases. Biology. 10 (9): 927.
9. Laing E. D., Sterling S. L., Weir D. L., Beauregard C. R., Smith I. L., Larsen S. E., Wang L. -F., Snow A. L., Schaefer B. C., Broder C. C. (2019). Enhanced Autophagy Contributes to Reduced Viral Infection in Black Flying Fox Cells. Viruses, 11, 260.
10. Jacquet S., Pons J.-B., De Bernardo A., Ngoubangoye B., Cosset F.-L., Régis C., Etienne L., Pontier D. (2019). Evolution of hepatitis B virus receptor NTCP reveals differential pathogenicity’s and species specificities of hepadnaviruses in primates, rodents, and bats. J Virol. 93: e 01738-18.
11. Subudhi S., Rapin N., Misra V. (2019). Immune System Modulation and Viral Persistence in Bats: Understanding Viral Spillover. Viruses, 11, 192.
12. Koh J., Itahana Y., Mendenhall I. H., Low D., Soh E. X. Y., Guo A. K., Chionh Y. T., Wang L.-F., Itahana K. (2019). ABCB1 protects bat cells from DNA damage induced by genotoxic compounds. Nature Communications 10: 2820.
13. Szentiványi T., Christe P., Glaizot O. (2019). Bat Flies and Their Microparasites: Current Knowledge and Distribution. Front. Vet. Sci. 6: 115.
14. O’Shea T. J., Cryan P. M., Cunningham A. A., Fooks A. R., Hayman D. T. S., Luis A. D., Peel A. J., Plowright R. K., Wood J. L. N. (2014). Bat Flight and Zoonotic Viruses. Emerging Infectious Diseases Vol. 20, No. 5.
15. Munshi-South J., Wilkinson J. S. (2010). Bats and birds: Exceptional longevity despite high metabolic rates. Ageing Research Reviews 9, 12–19.
16. Kuzmin I. V., Bozick B., Guagliardo S. A., Kunkel R., Shak J. R., Tong S., Rupprecht C. E. (2011). Bats, emerging infectious diseases, and the rabies paradigm revisited. Emerging Health Threats Journal, 4: 7159.
17. Caron A., Bourgarel M., Cappelle J., Liégeois F., De Nys H. M., Roger F. (2018). Ebola Virus Maintenance: If Not (Only) Bats, What Else? Viruses, 10, 549.
18. Foley N. M., Hughes G. M., Huang Z., Clarke M., Jebb D., Whelan C. V., Petit E. J., Touzalin F., Farcy O., Jones G., Ransome R. D., Kacprzyk J., O’Connell M. J., Kerth G., Rebelo H., Rodrigues L., Puechmaille S. J., Teeling E.C. (2018). Growing old, yet staying young: The role of telomeres in bats’ exceptional longevity. Sci. Adv.4.
19. Chionh Y. T., Cui J., Koh J., Mendenhall I. H., Ng J. H. J., Low D., Itahana K., Irving A. T., Wang L.-F. (2019). High basal heat-shock protein expression in bats confers resistance to cellular heat/oxidative stress. Cell Stress and Chaperones 24: 835–849.
20. Schountz T., Baker M. L., Butler J., Munster V. (2017). Immunological Control of Viral Infections in Bats and the Emergence of Viruses Highly Pathogenic to Humans. Front. Immunol. 8: 1098.
21. Guy C., Thiagavel J., Mideo N., Ratcliffe J. M. (2019). Phylogeny matters: revisiting ‘a comparison of bats and rodents as reservoirs of zoonotic viruses’. R. Soc. open sci.6: 181182.
22. Jebb D., Foley N. M., Whelan C. V., Touzalin F., Puechmaille S. J., Teeling E. C. (2018). Population level mitogenomics of long-lived bats reveals dynamic heteroplasmy and challenges the Free Radical Theory of Ageing. Nature. Scientific Report 8: 13634.
23. WynneJ.W., Shiell B. J., Marsh G. A., Boyd V., Harper J. A., Heesom K., Monaghan P., Zhou P., Payne J., Klein R., Todd S., Mok L., Green D., Bingham J., Tachedjian M., Baker M. L., Matthews D., Wang L.-F. (2014). Proteomics informed by transcriptomics reveals Hendra virus sensitizes bat cells to TRAIL-mediated apoptosis. Genome Biology 15: 532.
24. Hoelzer M., Schoen M., Wulle J., Mueller M. A., Drosten C., Marz M., Weber F. (2019). Virus and Interferon Alpha-Induced Transcriptomes of Cells from the Microbat Myotis daubentonii. iScience, 27: 647–661.
25. Irving A.T., Ahn M., Goh G., Anderson D. E., Wang L.-F. (2021). Lessons from the host defenses of bats, a unique viral reservoir. Nature. Vol. 589 (7842): 363–370.
Глава II. Минеральные ячейки в нуклеиновой обкладке: в начале была энергия
Отыщи всему начало, и ты многое поймешь.
Козьма ПрутковМитохондрии. Первый шаг к разгадке
Без митохондрий нет жизни животных, растений и грибов, но и ненасильственная смерть многоклеточных организмов неотрывно связана с этими удивительными структурами. Они – своеобразная альфа и омега для всех эукариот. Если каждая ныне живущая бактерия – это по сути клон древнейшей бактерии, жившей 3,5 миллиарда лет назад, просто с накопленной за это время гигантской суммой ошибок репликации при простом делении пополам и огромной сумятицей, вносимой горизонтальным переносом генов пару раз на тысячи поколений, то практически каждый эукариотический организм, даже одноклеточный, всегда приблизительно наполовину новый организм по отношению к своему предку, хоть и не всегда непосредственному (почкование или простое деление пополам, например, может служить таким пропуском предка). И именно митохондрия сделала из своего симбиоза с древней археей ту эукариотическую клетку, ту выдающуюся эволюционирующую машину, которая привела к возникновению животных, растений и всех пяти распознаваемых на сегодняшний день супергрупп эукариот. Ту эукариотическую клетку с ее врожденными противоречиями, проявляющимися или в виде типичных сбоев в ее функционировании, или в виде иных проблем, например с соседями, такими же, как она, эукариотами, или с совсем непохожими на нее вирусами и бактериями. То есть с врожденной предрасположенностью к болезням и, в конце концов, к смерти. Поэтому в отношении роли митохондрий для здоровья человека предлагается рассмотреть для начала только два самых важных аспекта, сообразующихся с центральной идеей данной книги: как появилась электрон-транспортная цепь (ЭТЦ) – ключевое событие для возникновения жизни[2] и самый важный энергетический механизм в митохондриях, и как правильно организованный энергетический метаболизм клетки определяет ее гомеостаз и адаптационные способности организма. Если рассматривать эволюцию именно энергетического метаболизма от самых истоков зарождения жизни, то мы по прямой линии скорее наследники бактерий-предков митохондрий, чем архей, приютивших их хозяев.
К горячим источникам
По крутым, покрытым облаками склонам узкого ущелья к мелкой горной речке сползают сине-зеленые волны высоких сосен. Иногда в утреннем тумане возникает просвет, и, если вглядеться, на крутом правом берегу проступают контуры полуразрушенного каменного строения метров двадцать высотой. Я сижу на противоположном берегу, где спуск к реке более пологий, на ступенях недавно отреставрированного великокняжеского дворца XIX века в мавританском стиле. Километрах в двух отсюда, уже на левом берегу, есть похожие, но совсем малозаметные за городской застройкой развалины второй каменной башни. Это останки «парных» крепостей легендарных братьев Петре и Гогиа Авалишвили – феодалов с явными разбойничьими наклонностями, постоянно враждовавших друг с другом, но изредка и замирявшихся. Крепость Петре – Петрес-цихе – была главной, но только вместе со второй – Гогиас-цихе – обретала полноценное стратегическое значение: крепости охраняли древнюю переправу через Куру, по которой часто шли в центральную Грузию захватчики с юга.
В голову приходит мысль, что здесь может быть еще один вариант разгадки происхождения названия этого места. Тысяча триста лет назад сюда пришли арабы. Возможно, укрепления на этих местах существовали и до [без]башенных братьев Авалишвили. «Две башни» – по-арабски «боржан» («боржан» одно слово, без числительного, так как семитские языки – из ныне существующих иврит и арабский – сохранили уникальную форму двойственного числа существительных, практически утраченную в индоевропейских языках). В местном картлийском наречии «боржан» – «боржани» за века легко могли превратиться в «Боржоми». Старые источники «Боржоми» находятся как раз неподалеку, с другой стороны Куры, где к стремительной горной реке торопливо подбирается не то маленькая речка, не то большой ручей Боржомка.
Ранним утром возле павильона «Боржоми» еще совсем мало курортников, и вода из крана льется не полной струей, а прерывистой маленькой струйкой, но все равно чувствуется устойчивый запах сероводорода. Сто семьдесят лет назад здесь была яма, заполненная слегка замутненной, но сильно пахнущей водой. Со дна ямы била мощная горячая струя. Сильно щелочная вода источника зарождается в нескольких километрах под землей, в толщах вулканической породы, где еще незастывшая магма подбирается исключительно близко к земной поверхности. Считается, что часть этой воды имеет ювенильное происхождение, то есть образуется в ходе прямой реакции выделяющихся из магматических масс кислорода и водорода. Хотя это (и не только) делает боржомскую воду весьма особенной, но на самом деле щелочных гидрокарбонатных источников иного происхождения на Земле всегда было относительно много как на поверхности, так и в морской глубине или на границе воды и суши. Вполне возможно, щелочные источники могли возникнуть и на дне или берегу соленого озера, похожего на нынешнее Мертвое море, расположенное, кстати, неподалеку от другого места с таким же двойственным названием – Ерушалаим (Иерусалим). Именно двойственность, точнее, дополнительность двух базовых сущностей, лежащая в основе базовых биологических форм – многоклеточного организма, эукариотической клетки, просто первой клетки или первичного протоклеточного состояния, обладающего всеми признаками живого организма, и многих других, – составляет второй основной лейтмотив данной книги, наряду с упомянутой во вступлении сквозной фрактальностью биологических форм движения материи.
Горячие щелочные гидротермальные подводные источники все чаще называются наиболее вероятным местом возникновения жизни, по крайней мере в рамках «первично-метаболических» теорий.
Гюнтер Вехтерсхойзер в начале 80-х указал на принципиальную возможность образования в таких источниках органических молекул путем восстановления углекислого газа при участии восстановителей – сероводорода или водорода – на поверхности сульфидов железа, сгруппированных в железосерные кластеры. Очень важно подчеркнуть, что невероятно похожие железосерные кластеры являются коферментами, то есть «рабочими частями» важнейших ферментов энергетического и дыхательного метаболизма у всех живых организмов. Если Гюнтера Вехтерсхойзера вдохновляли завораживающие картины гидротермальных «черных курильщиков», перегретых магмой водных потоков, перенасыщенными сероводородами, сульфидами, водородом, вырывающимися на двух-трехкилометровой глубине черными клубами в толщу океана, то Майк Рассел и Билл Мартин перевели фокус внимания на горячие щелочные гидрокарбонатные источники, часто располагающиеся на гораздо меньшей глубине или вообще на суше. Эти источники также содержат водород, сероводород и железо, правда, как правило, в гораздо меньшей концентрации, меньше простейших соединений углерода, и имеют меньшую температуру, так как образуются не в результате взаимодействия с магмой, а в результате химического взаимодействия с мантийными породами («Боржоми», кстати, и в этом смысле может быть исключением: как упоминалось выше, он может быть в значительной степени ювенильной водой, результатом прямой реакции кислорода и водорода магмы, причем водород участвует в явном переизбытке; также в «Боржоми», хоть и в крайне незначительных количествах, обнаруживаются разнообразные простые одно- и двууглеродные соединения, в том числе азотно-углеродные). Как существующие ныне подводные щелочные гидротермальные источники, так, очевидно, и древние часто имели строение проводящей их породы в виде микропористой губки, с ячеистостью от нескольких сантиметров до долей миллиметра. Такое строение позволяет поддерживать полупроницаемый барьер между двумя видами сред: условно наружной, кислой и богатой натрием, и внутренней, щелочной. Одновременно малые размеры ячеек обеспечивают возможность создания внутри них достаточно высоких концентраций углекислоты, сероводорода и(или) водорода, которые в присутствии каталитических железосерных кластеров и в условиях повышенной температуры и высокого давления могут образовывать примитивные органические молекулы. Необходимый приток энергии самоорганизуется за счет разности потенциалов, созданной полупроницаемыми неорганическими (на тот момент) барьерами. Собственно, это практически уже две из трех форм биологически свободно конвертируемых форм энергии, согласно первому закону биоэнергетики академика РАН Владимира Скулачева (первая и вторая – это натриевый и протонный (водородный) потенциалы; третья форма – АТФ). В более широком смысле – это возникновение и, что более важно, стабилизация химической неуравновешенности и молекулярной упорядоченности, что в терминах термодинамики можно назвать локальным уменьшением энтропии[3]. Обобщая, можно сказать, что наличие большого количества легкодоступной энергии соответствует низкоэнтропийному состоянию системы, и, напротив, дефицит легкодоступной энергии – высокоэнтропийному.
Если к горячим источникам добавить цикличность
В последние годы исследовательская группа Тары Джокич, Дэвида Димера и Мартина ван Кранендонка (Tara Jokic, David Deamer and Martin van Kranendonk, 2017) активно разрабатывает альтернативную теорию наиболее вероятного места и механизма происхождения жизни. Они располагают это сакральное место в окрестностях систем древних наземных вулканических водоемов, похожих на современные гейзерные поля Йеллоустоуна или Камчатки, но обладавших цикличностью высыхания-гелеобразования-увлажнения. Подобная цикличность позволяет образовываться многослойным протоорганическим формированиям, предшественникам органических полупроницаемых мембран – первоначально, в сухую фазу, плоским, и изредка во влажную фазу, со сферическими отпочкованиями. Однако энергетический баланс протоклеточных структур в этом случае с трудом покрывается простыми неорганическими источниками энергии, например полифосфатами. Принципиально эта теория не отвергает роль щелочных гидротермальных источников: на примере Боржоми мы видим, что такие источники вполне могут быть наземными; более того, в историческом плане они могут быть наземно-приморскими: та же местность Боржоми еще в позднем миоцене, возможно, представляла собой прибрежную зону деградирующего океана Тетис с разворачивающейся вулканической деятельностью. Вероятность подобных сочетаний в эоархее, предполагаемой эпохе возникновения настоящих жизненных форм, в общем-то тоже не исключена. В подобных случаях цикличность может включать два типа влажных фаз (или серий фаз): щелочную гидротермальную и кислую морскую (приливную?). В любом случае крайне маловероятно, что даже в «чистой» теории щелочных гидротермальных источников линия разграничения щелочного и кислого потоков будет стабильна; более вероятно, что в части микроячеистой породы с большей проницаемостью попеременно меняется рН среды, а в части микроячеек с меньшей проницаемостью рН более стабильно, но меняется их окружение. Возможная «сухая» фаза способствует дополнительной концентрации органических молекул на первичных минеральных мембранах, дополнительно к феноменам термофорезаи компартментализации (то есть концентрирования в пограничных сегментах за счет тепловых градиентов и тепловых конвекционных потоков во множестве полупроницаемых разграничений). В принципе, как показано исследовательской группой под руководством Джулиана Тэннера и Андерсона Шума из Университета Гонконга на примере термодинамики «высыхающих капель», компартментализации в протобиологических структурах может способствовать даже процесс частичного испарения в двухфазовых водных системах (Guo W. et al., 2021).
В последующем неорганическая основа может почти полностью «вымываться» или механически, или кислотным воздействием, подобно вытравливанию кислотой элементной платы, оставляя функциональные каталитические фрагменты – те же железосерные кластеры. С другой стороны, «чистая» теория щелочных гидротермальных источников «выталкивает» наиболее вероятных прародителей биологической изменчивости и наследственности – это неустойчивые в щелочной среде полимеры рибонуклеиновой кислоты (РНК) – на кислую сторону первичных полупроницаемых мембран, где этой теорией не предполагается (хотя и не исключается) ячеистая неорганическая мембранная структура (компартментализация). В комбинированном циклическом варианте с двумя – кислой и щелочной – влажными фазами фрагменты РНК могут быть заключены в ячейку с кислой средой внутри, находящуюся в щелочном окружении. Более того, подобным образом могут организовываться вторичные и третичные матрешкообразные ячеистые формирования, где мелкие кислые пузырьки в окружении более крупных щелочных конгломератов, оказывающиеся внутри крупных кислых кластеров, частично окруженных щелочной средой и так далее (рис. 2–6).
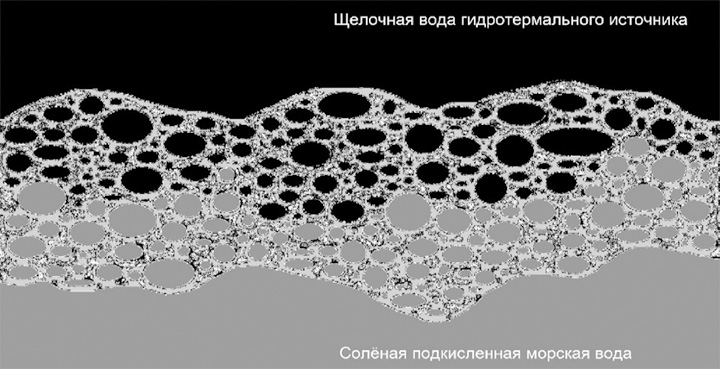
Рис. 2. Микропористые ячейки, проводящие щелочную воду гидротермального источника, частично заполнены также подкисленной соленой морской водой
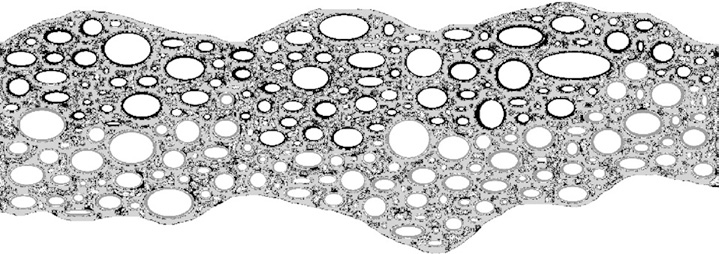
Рис. 3. Сухая фаза: минеральные и органические осадки из щелочной воды источников и соленой морской воды оставили отложения на стенках микроячеек (отмечены черным и серым цветом соответственно)

