


Полная версия
Collins New Naturalist Library
Phylum Mollusca
There are many cavernicolous Gastropoda within the families Auriculidae, Zonitidae, Subulinidae, Enidae and Valloniidae (terrestrial species), and Hydrobiidae (aquatic species). The Bivalvia include cavernicolous species of Pisidium and Congeria in Europe and Japan.
Phylum Onychophora
The South African species Peripatopsis alba is known only from caves.
Phylum Arthropoda
Subphylum Crustacea
Class Remipedia: Recently discovered in submarine caves in the Bahamas and Canary Islands, these actively swimming predators resemble aquatic centipedes.
Class Ostracoda: There are many groundwater species, some of which also occur in larger caves.
Class Copepoda: Many cyclopoids and harpacticoids occur in caves and interstitial groundwaters worldwide.
Class Malacostraca: The monospecific order Spelaeogriphacea is so far known only from caves in South Africa. Thermosbaenacea and Mysidacea are widespread in submarine caves and brackish groundwaters and a few species have made the evolutionary shift into freshwater caves. Isopoda are one of the best-represented orders in the subterranean world, with hundreds of cavernicoles described within the Asellidae and Oniscoidea. The Amphipoda, notably the Gammaridae, are equally well represented. Cavernicolous decapods such as crayfishes, galatheids, crabs and river prawns are particularly widespread in the tropics. Groundwater-inhabiting members of this class often belong to ancient lineages and the distributions of related taxa provide important evidence in reconstructing the history of the planet.
Subphylum Uniramia
Class Diplopoda and Pauropoda: Worldwide there are hundreds of species of cavernicolous millipedes, and particularly of Polydesmoidea. They often show relictual distributions which mirror past configurations of the earth’s crustal plates, long since redistributed by seafloor spreading and continental drift. A few Pauropoda are known from caves.
Classes Chilopoda and Symphyla: Cavernicolous Scolopendromorphs and Lithobiomorphs are known from European caves. In the tropics there are also cave-evolved Scutigerids. The Symphyla live in the soil and look like cave animals, but most feed on living plant roots and so are excluded from deep cave habitats.
Class Insecta: The Collembola and Diplura are primitive, wingless, ground-dwelling insects which require a high humidity. They include a large number of cave-evolved species worldwide. Cavernicolous Blattodea are mostly confined to the tropics. Within the Orthoptera, the Rhaphidophoridae or camel crickets contain a number of conspicuous cavernicoles with a worldwide distribution and a few cavernicolous gryllids are found in the tropics. There are cave-evolved Dermaptera, Psocoptera, Hemiptera, Trichoptera, Lepidoptera and Diptera. Not surprisingly, the majority of cavernicolous insects belong to the largest order and the most successful group of animals on earth, the beetles (Coleoptera). At least 22 families contain cavernicoles, which reach their greatest diversity and specialization in the Trechidae and Leiodidae.
Subphylum Chelicerata
Class Arachnida: A few cavernicolous Scorpiones, Uropygi, Amblypygi, Schizomida, Ricinulei and Palpigradi are known from tropical regions. Pseudoscorpions are well represented in caves, with over 300 cavernicolous species, many of which are giants among their kind, with very long legs and claws. The Opiliones also contain cavernicoles, most notably in the families Phalangodidae, Travuniidae and Ischyropsalidae. The terrestrial mites (Acari) most frequently found in caves are tiny Gamasides within the families Parasitidae, Rhagidiidae and Eupodidae and there are many cavernicolous water mites in the families Hydrachnellae and Porohalacaridae. Spiders (Araneae) dominate the predator niches in most tropical caves and include hundreds of cavernicolous species worldwide. In temperate regions, the most specialized cavernicoles belong to the primitive families Dysderidae, Leptonetidae, Telemidae and Oonopidae, but there are also many specialized species of Linyphiidae, Erigonidae and Agelenidae.
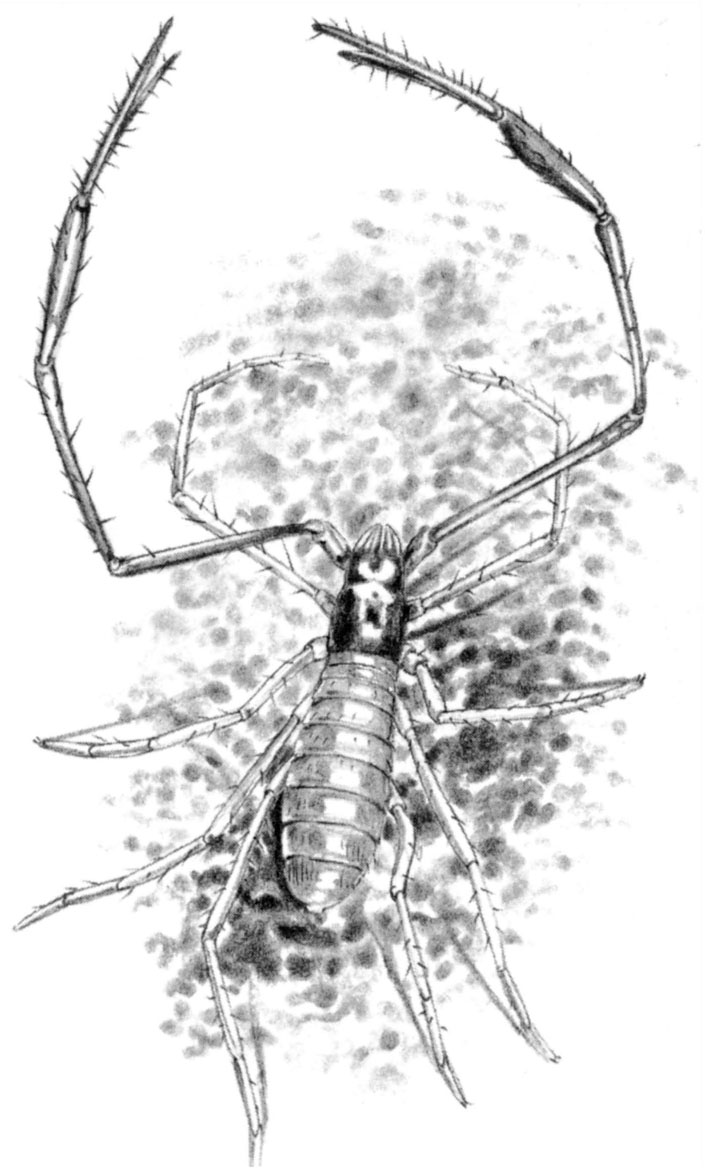
Fig. 2.3 Neobisium spelaeum, a giant cavernicolous pseudoscorpion which preys on the cave beetle Leptodirus hohenwarti (shown in Fig. 2.2)
Phyla Tardigrada and Bryozoa
A few species inhabit groundwaters.
Phylum Echinodermata
Various detritivorous brittle stars (Ophiuroidea) and sea cucumbers (Holothuroidea) frequently occur in submarine caves, but no strictly cavernicolous species are known.
Phylum Chordata
Class Teleostomi: Freshwater cavernicolous fishes are found mainly in the more arid regions of the world. To date around 60 more or less blind and depigmented species representing 8 orders and 13 families have been collected in freshwater and submarine caves, springs and groundwaters. The most speciose families are Cyprinidae, Gobiidae and Bythitidae, followed by Pimelodidae, Characidae, Cobitidae and Amblyopsidae.
Class Amphibia: 14 species of cavernicolous Urodela are known from North American caves and one from Europe. The latter, Proteus anguinus was the first cavernicole to be recognized as such. In common with species of the American genera Gyrinophilus, Eurycea, Typhlomolge and Haideotriton, Proteus retains larval gills in the adult stage, and is thus able to live permanently beneath the water table.
Class Reptilia: The colubrid snake Elaphe taeniura is a common inhabitant of caves in south-east Asia, from China to Borneo, where it preys on bats and swiftlets.
Class Aves: While no birds live permanently in caves, swiftlets of the genus Aerodramus (Apodidae) and the oilbird Steatornis caripensis (Caprimulgidae) are dependent on caves as nesting sites and are capable of navigating long distances underground and in total darkness by echolocation using ultrasonic clicks.
Class Mammalia: It is possible that a South American mouse Heteromys anomalus and one or two species of tropical shrews in the genus Crocidura may establish permanent cave populations. Many bats (e.g. Tadarida spp.) are dependent on caves for shelter, but none are cavernicolous.
In summary, although green plants are largely absent from caves, cavernicolous species are found within most of the major classes of animals, and are particularly common among the Crustacea, insects, spiders and millipedes.
Caves in limestone
Of several cavernous rocks in Britain and Ireland, one above all others provides extensive integrated cave systems of a size which allows the biologist to study the life they contain with relative ease. It is the Carboniferous Limestone.
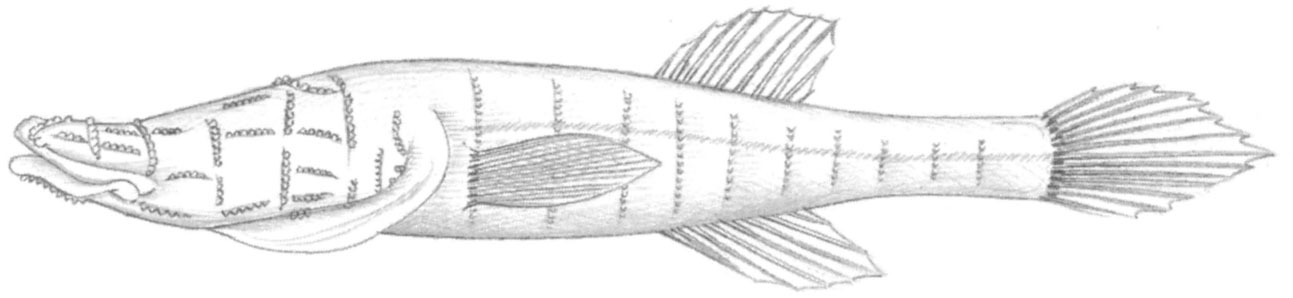
Fig. 2.4 The blind North American cave fish Speleoplatyrhinus poulsoni has an extraordinarily developed lateral line system, with correspondingly enlarged brain parts for processing tactile and positional information.
The cave-bearing limestones are concentrated in six major patches: in County Fermanagh (the largest, but least well prospected area), the Northern Pennines, Derbyshire’s Peak District, County Clare, Somerset’s Mendip Hills and South Wales. In the Northern Pennines alone, there are 1800 or more documented cave entrances and around 350 km of explored and mapped cave passages – while current estimates give the total length of mapped passages in Britain and Ireland at somewhere around 800 km. Cave exploration is still in a very active phase in these islands, and significant new discoveries are still being made. For example, recent explorations in Ogof Daren Cilau beneath Mynydd Llangattwg in South Wales have yielded over 20 km of new discoveries in less than two years, including the largest passages yet found in Britain.
By any standards, 800 kilometers of open cave passage represents a significant habitat, worthy of the attention of naturalists – yet passages of explorable size must form but a tiny proportion of the total cave habitat, the vast majority being of mesocavernous dimensions. In the absence of data, I would guess that the habitable surface area within the mesocaverns of limestone terrains must run to at least two or three orders of magnitude more than that within explored caves.
So what is it which makes limestones so spectacularly cavernous? To understand the process in which caves are formed, we must begin by examining the origins, nature and structure of the limestone rock itself. Limestone is a sedimentary rock, that is, it began life as suspended particles in an ancient sea, gradually settling to the ocean floor millimetre by millimetre over millions of years. During this inconceivably long period, there were intervals when conditions changed enough to interrupt the steady downward rain of lime, allowing some other type of deposit to intervene briefly in the sequence of otherwise pure calcium carbonate. Aeons later, and now hardened to rock, these geological glitches have become ‘fossilized’ as bedding planes – horizons of weakness between the solid layers of limestone.
The Carboniferous Limestones of the British Isles were laid down somewhere in the tropical seas of the southern hemisphere. Pushed along on a northward-drifting chunk of continental crust, they have had a bumpy ride. Some, like those of the Yorksire Dales, have survived their 340 million year journey the right way up, though somewhat fragmented by massive vertical faults. Others have fared less well. The Mendip limestones lie like a wrecked car, buckled and perched at a steep angle, so that the bedding planes dip downhill at an average gradient of 50° or so. In all cases, the rough ride has produced vertical stress cracks, called joints, which link with faults and with the original bedding weaknesses of the limestone to form a boxwork of crevices reaching from the highest hilltop to beneath the deepest valley.
Limestone is a strong rock and so frequently forms upland regions. Solid limestone is impervious to water, but water is able to flow through the cracks within it. It is these cracks which are the key to understanding the origins of caves. Limestone caves form principally by means of a simple chemical reaction in which hydrogen ions from groundwaters, acidified with dissolved carbon dioxide, act on the relatively insoluble carbonate ions in the limestone to produce soluble bicarbonate 10ns which are then flushed away. The reaction renders the limestone 25 times more soluble than it would be in pure water and the result is holes.
Some of the carbon dioxide in groundwaters is collected by raindrops falling through the atmosphere, and some from the breakdown of organic material picked up as the rain then trickles down through the soil. Immediately beneath the soil, the weathered surface layers of rock are more fractured than those at a greater depth and the acidified, aggressive soil waters have their maximum impact here – so that at any one time, up to 15% of the volume in the top three metres of limestone may be occupied by air-, or water-filled spaces (Stearns, 1977 – figures for Central Tennessee, USA). These mesocaverns act like the guttering beneath the roof of a house – collecting soil water and quickly conveying it to natural drainpipes, often developed on the intersections of major joint fractures, or steeply inclined bedding planes.
The flow of water downwards into the limestone carries with it sediments which contain particulate organic material, dissolved organic acids and microorganisms (bacteria and fungi). Decomposition continues way below the soil in the cracks and crevices of the limestone. There, to paraphrase Hoover’s famous advertising slogan, groundwater micro-organisms ‘eat-as-they-seep-as-they-clean’, mopping up the organic impurities and excreting CO2 – in effect arming their liquid medium with chemical teeth. In the larger conduits it may take up to 50 days and several kilometres of flow before the bacteria finish mopping up the water-borne food, and even longer and further before the chemical aggression of the cave water is finally spent. All this time, limestone is being steadily corroded, and the cracks along which the water travels widened, resulting in the slow opening of a complex drainage network reaching deep below the water table.
The initial pattern of flow within the flooded cracks is dictated by the structural geology of the limestone and the shape of the land surface. Between them, these two factors determine where water will escape from the rocks as a spring, and lay down the blueprint for the caves to come. If the rock strata lie horizontally, as they do in the Yorkshire Dales, water is forced to follow a rectangular course down vertical joints and along short sections of horizontal bedding, producing the characteristic stepped profile of a ‘pothole’ system. If the strata are inclined, as in the Mendip Hills, drainage will alternate between short joint-controlled shafts and longer bedding-plane slopes, producing caves with a steep profile, but few vertical sections.
At or below the water table, the course taken by percolation water is determined by the direction of the hydrological gradient between where the water goes into the permanently-flooded system of cracks known as the ‘phreas’ and where it comes out again as a spring. Within the phreas, water is free to follow along, down or up the 3-D maze of cracks to produce the smoothest possible overall flow. Where the rock beds lie horizontally the smoothest profile may be along just one bed, producing a horizontal cave which may run for several kilometers (Yorkshire has many such systems). Where the rocks dip steeply in a direction which does not coincide with the hydrologically-determined direction of flow, the groundwater may be forced into a series of vertical z-bends, down joints and up the bedding, to tack its way to the spring (a classic example is Wookey Hole in the Mendip Hills). Where the limestone beds are trapped in a syncline or U-bend beneath impermeable rocks, water may be forced to travel down to great depths following the configuration of the rock strata. Chinese geomorphologist Yuan Daoxian has recently reported the discovery of a substantial cave at a depth of 2900 m below the water table in the Sichuan Basin in China. When perforated by drilling, the hole gushed water. Caves at this depth are, however, extremely rare and probably have little or no biological significance.
As the profile of the cave takes shape, under the twin controls of hydrology and geology, the ‘best route’ is inevitably favoured with a greater rate of flow, which promotes more rapid corrosion of its boundary walls – which in turn results in a still greater flow capacity. In this way, the initially diffuse drainage within the limestone is gradually simplified with increasing time (and depth) into a pattern of coalescing collectors of increasing diameter. Over the aeons, the vagaries of geology and hydrology may conspire to favour one particular route in preference over all others, opening it out to form a major trunk conduit which drains the entire subterranean aquifer to its spring, or ‘resurgence’.
I have suggested so far that cracks in the limestone can be enlarged to form mesocavernous conduits by corrosive water trickling gently down towards the water table under the influence of gravity, or by slow creeping but aggressive groundwaters draining towards a resurgence. However, chemical solution is not the full story – once the cave becomes large enough to be an efficient drain, fast-moving waters can carry sharp-edged grit, adding physical claws to chemical teeth. Grains of quartz (a mineral much harder than calcite) provide a particularly effective scouring agent, and are easily collected by streams running over the Millstone Grit which butts onto much of our cavernous limestone. Where such a stream is captured by a well-developed Yorkshire joint, the outcome is a vertical pothole; on Mendip, an inclined swallet cave results.
Erosion of the river valleys which drain a limestone block may lower the water table within it, so that a conduit formed within the flooded phreas eventually becomes emptied of water and filled with air. One such system, GB Cave in the Mendip Hills, has received intensive study in recent years – in turn by Drs Tim Atkinson, Pete Smart and Hans Friederich of Bristol University. Their work has demonstrated the extent and importance of the system of mesocavernous conduits which overlies and feeds water and sediment into GB Cave. The conduits can be divided into three types. First there is the dense network of ‘subcutaneous’ cracks in the top three metres of rock beneath the soil. These have a large total volume and drain rapidly into the underlying cave, via mesocavernous collectors. They operate intermittently for up to two days following rain, but do not store a great deal of water. Two other types of inlets feed into the cave. They are small-diameter seeps, which stay full of water all the time and show little variation in flow discharge (flow being constrained by their small diameter); and vadose flows, which have higher and more variable discharges than do the seeps. Smart and Friedrich report that inlets are more frequent into shallower cave passages, and less frequent at depth, but do not give estimates of the porosity represented by these inlets, for which we have to consult Stearns on his Tennessee karst. Thus, at 10 m depth, the initially high porosity within the zone of subcutaneous flow (15%) may have fallen to about 1.5%, with a mainly downward flow via mesocavernous shafts and vadose cracks. At 30 m depth, porosity is down to 0.002%, and flow (at least in massively-bedded limestones) is increasingly confined within macrocavernous collector passages, fed by a converging network of smaller collectors of mesocavernous size. Thus, the main concentration of cave habitats (of mesocavernous dimensions) lies within the top three metres of bedrock.
Fig. 2.5 The main streamway in Poulnagollum Pot, Co. Clare – a classic vadose passage cut down through the limestone beds by an allogenic stream flowing off the eastern flank of Slieve Elva (compare with Fig. 2.1). (Chris Howes)
Phreatically-formed passages can be recognized by their characteristic circular or at least symmetrical cross-section, produced by equal corrosion of the walls, ceiling and floor (a classic example is Peak Cavern in Derbyshire). As soon as they develop an air surface, corrosion and erosion become channelled downwards and the cave stream begins to cut a trench into the floor, as is seen in the Ogof Ffynnon Ddu streamway in South Wales. This may eventually result in a canyon-shaped passage many metres deep.
As the cave matures, the enlargement of joints and bedding planes may weaken whole blocks of the ceiling and floor, resulting in their collapse. Where the cave contains an aggressive stream, such material may be removed as it falls, creating a chamber whose size is limited only by the structural strength of the overlying rock. Given enough time, the roof will eventually fail, opening the cave to the sky; if the collapsed blocks are not removed by water, the cave fills up with rubble.
The same chemical process which resulted in the removal by solution of limestone in cracks close to the surface, can go into reverse when acidified, lime-laden waters drip through cracks into an underground passage full of fresh air. As CO2 in the hanging water droplet diffuses out into the air of the cave, the resulting transformation of bicarbonate to carbonate ions forces lime to precipitate from solution. The drip falls, a rim of lime remains on the cave ceiling, and hey presto, a few thousand years later the cave is festooned with sparkling stalactites. Given an aeon more, these may completely fill up and block the passage.
Caves may also become filled with insoluble sediments from outside. This is frequently the case in Britain, where sediments enter the cave by one of three main mechanisms: large masses of unsorted clay-and-rubble have slumped into caves as a half-frozen mush during the glacial advances of the Pleistocene; sands and gravels are washed in by cave streams and dumped as the waters slow down in more gently graded stretches of cave; and fine mud is often trickled downwards by percolation water until it completely fills passages right up to the roof.
The worst ravages of collapse and sedimentation may be reversed by a little-known bio-chemical phenomenon known as ‘digging’. First noted in Mendip caves, it appears to be caused by large sweaty men in overalls, armed with plastic buckets, and generally takes place at weekends in the pursuit of new explorations and discoveries underground. The chemical component of the process consists in a sparing application of ‘Dr Nobel’s Linctus’ to otherwise immovable blockages, with sometimes impressive results. We shall consider the conservation implications of digging and blasting in caves in a later chapter.
Types of cave habitat
In the previous section, we traced the process of cave formation in limestones by the gradual chemical enlargement of bedding cracks and joints, their subsequent drainage, further enlargement, collapse and infilling by sediment, breakdown and speleothems. In the course of this ‘life cycle’, limestone caves present a series of distinct habitats which are each exploited by a characteristic biota.
In recent years, similar biotas have been found in equivalent habitats in a wide range of other rocks (lava, gypsum, mudstone, shale, chert, breccia, tuff, rhyolite, diorite, quartzitic sandstone, etc) and sediments. This has led to a plethora of exotic technical terms in the literature – it seems that any biologist who finds a new habitat wants to give it an impressive, polysllabic label, an urge doubtless born of years of grappling with latin species names. My favourite examples include ‘parafluvial nappes’ (as worn by water-babies?), ‘hypotelminorheic biotope’ and ‘the petrimadicolous biocoenose’ (etymologically-inclined readers may enjoy trying to sort out these little beauties). In the section which follows, I have attempted to keep things simple by stressing the the similarities between habitats, rather than the differences between them.
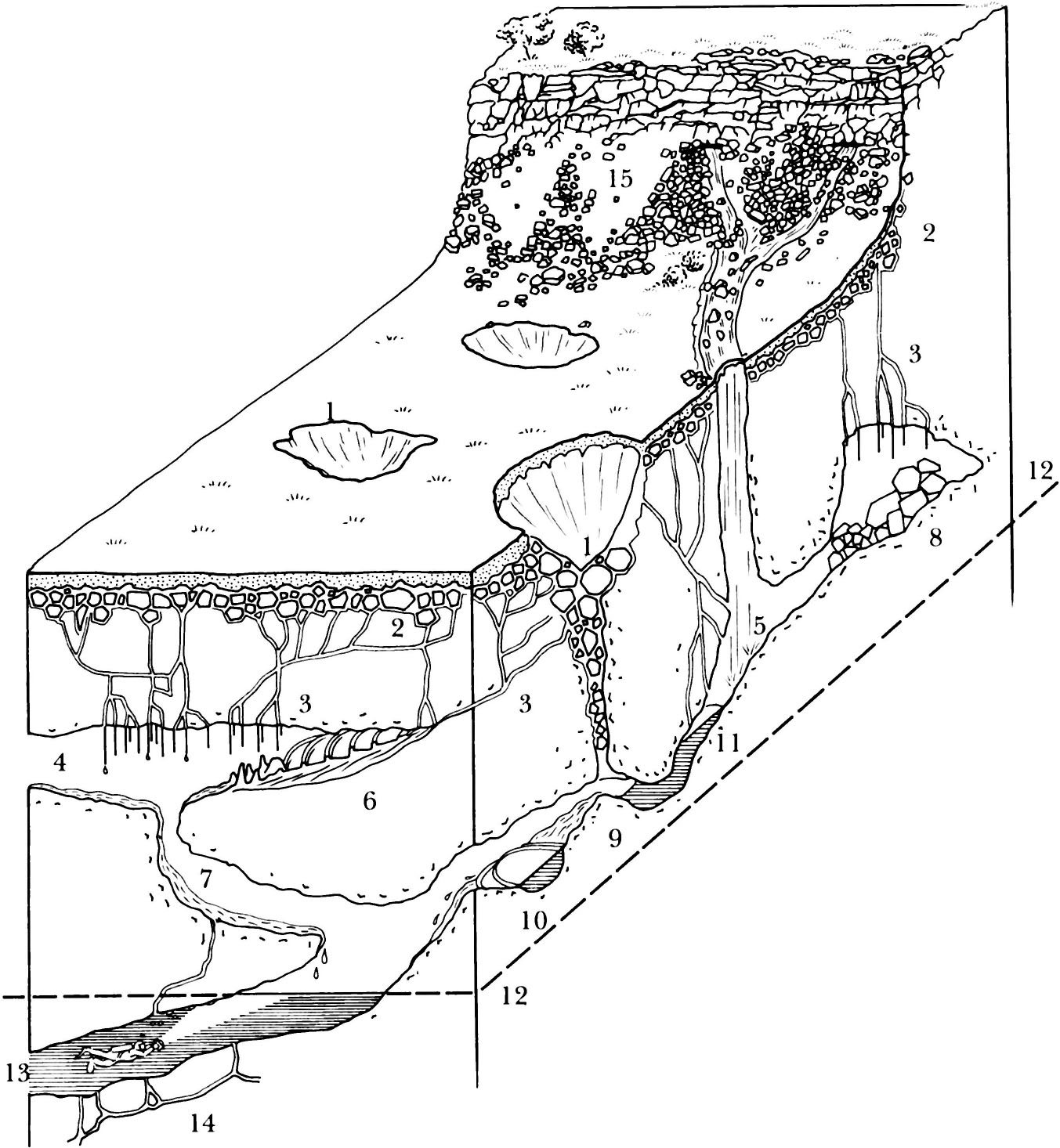
Fig. 2.6 Cave habitats and structural features in a limestone upland.
1. Dolines or shakeholes 2. SUC or subcutaneous zone 3. Mesocavernous seeps and conduits 4. Air-filled macrocavern or dry cave passage 5. Vadose macrocavern or wet cave passage 6. Rimstone gour pools 7. Cave sediment 8. Breakdown blocks 9,10. Riffle and pool in cave system 11. Perched siphon or sump 12. Water table 13. Water-filled macrocavern or sump in the phreas below water table 14. Water-filled mesocaverns in the phreas 15. Talus, below a scar or limestone cliff.
Air-filled macrocaverns (dry caves)
The longest, biggest and, by any criteria, the best macrocaverns are those developed in limestone. The world’s longest cave is the Mammoth Cave system in Kentucky at over 540 kilometres, while the deepest is the Réseau Jean Bernard in the French Pyrenees at over 1600 m deep. Our longest cave, the Ease Gill system in Yorkshire, 70 km long, can manage only 15th place in the world ratings. The ages of caves vary considerably. Recent estimates suggest that certain limestone cave passages in North America could have formed 10 million years ago, while some tropical limestone caves are estimated to be only around 100,000 years old.

