


Полная версия
Collins New Naturalist Library
2
The Cave Habitat
What is a cave?
Put this question to any wetsuit-clad, hard-hatted individual found walking across the Mendip Hills, Yorkshire Dales or the shining limestone pavements of the Burren, and you will discover that a cave is a naturally-formed hole in limestone which is large enough to be explored by a caver.
Ask the same caver what he or she has noticed in the way of living creatures in caves, and the answer may well be “not a lot.” It will perhaps surprise most cavers (and naturalists) to learn that over a hundred species of invertebrate animal have been recorded as maintaining permanent populations in cave habitats in Britain and Ireland, plus another score or so species of creatures such as bats and moths which use caves as a regular part-time shelter. The cryptic community to which these creatures belong remains largely undetected by cavers because it generally avoids the relatively large tunnels and booming chambers, the glistening calcite draperies and crashing waterfalls which so captivate the human cave enthusiast, preferring instead the cosy confines of smaller cracks and crevices. I hope in this book to shed light on at least a portion of this unsuspected world which lies beneath our feet, whether we live in Grassington or Glasgow, Lisdoonvarna or London, and to offer pointers to fellow amateur naturalists towards fruitful areas of investigation for the future.
To unravel the natural history of a cave, or indeed any habitat, we must try to perceive it as far as possible from the point of view of its inhabitants. It requires a considerable effort of imagination for such sight-dependent creatures as ourselves to grasp the essence of life in the dark and labyrinthine realm of the cavernicole. A little inspired speculation may be needed to find the ‘right’ questions to lead us to fresh insights into the mysterious world beneath us.
We might start by imagining what kinds of habitats could be accessible to the cavernicole and then consider which environmental characteristics of such places are likely to influence its choice of where to live. Almost at once we run into problems, for although a good deal is known about the environment in man-sized, air-filled limestone caves in Britain and Ireland, we know much less about the conditions within smaller cracks and crevices or in caves beneath the water-table, and less still about our submarine caves. Fortunately, such information is more readily available from other parts of the world. So in this chapter we will take an international approach to defining and classifying the cave environment, before turning in later chapters to a detailed consideration of what is known about our own cave fauna.
What then is a ‘cave’ as perceived by its inhabitants? The dictionary definition of “a natural underground chamber” gives us a less than helpful starting point, for why should we suppose that the cavernicole will distinguish between natural or man-made tunnels, or between subterranean and above-ground enclosed chambers, as long as the appropriate conditions of food supply and microclimate are present in the living space? Should we then include mines, adits, buried pipes, culverts, sewers, cellars, tombs, the London Underground System, or perhaps even houses and other enclosed buildings in our preliminary list of potential cave-habitats? To what extent should our definition specify the material bounding the cavity? Must a cave be rock-lined, or should we widen our brief to include animal burrows and other spaces present in soil, for surely it must be arbitrary to distinguish between an earthy burrow and a muddy hole of similar dimensions in rock?

Fig. 2.1 The main tunnel of Sleets Gill Cave in Wharfedale, Yorkshire Pennines – a classic phreatic tube, formed and enlarged by water filling the passage and so dissolving the limestone rock equally on all sides. (Chris Howes)
As it happens, soil faunas are very well documented, and while it seems that many animal groups are common to both soils and caves (and indeed to leaf-litter and the deep moss-carpets of tropical regions as well), the fauna of organically-rich topsoils is sufficiently distinct from that of most rock-space habitats to warrant a separate treatment. I shall therefore exclude soils forthwith from our definition of cave habitats (but see ‘Cave sediments’ under ‘Types of cave habitat’ later in this chapter). Similarly, the voids in other organic, living or once-living materials, such as wood or the guts or blood vessels of animals, have distinctive specialized faunas of their own, clearly distinguishable from those of habitats within inorganic materials – although some specialized xylophages, such as termites, and endoparasites, such as tapeworms or flukes, share certain morphological specializations (eyelessness, depigmentation) characteristic of the more specialized cavernicoles. When it comes to holes of human fabrication, most significant biological criteria must lead us to include them in our category of caves. That they have a very poor fauna in comparison with natural caves, is due less to their artificial nature than to their frequent isolation from sources of natural colonization and their often unfavourable microclimate.
Having narrowed our definition of the cave environment to ‘habitable voids bounded by walls of rock, or similar inorganic materials’, let us now consider the physical criteria which may determine their habitability: the presence or absence of light, physical space (the size of the hole), the medium filling the space (water or gas mix), the microclimate within the medium (the pattern of change in temperature, pH, etc. over time), and the nature and amount of available food.
Let us begin with the business of light, a variable of obvious biological significance. Beyond the limits of light penetration, the cavernicole will be obliged to rely on senses other than sight, and on foods other than green plants. Perpetual darkness is a characteristic of most rock void habitats anyway, so let us choose to define ‘the cave’ as a habitat entirely without natural illumination. This will substantially simplify our task, by excluding from the cave fauna a whole host of organisms which seek shelter in cave entrances, but also live in a wide range of other shady, sheltered habitats such as the woodland floor, or river gorges, or houses and other structures used by people. Later we will consider the illuminated portions of man-sized caves as a significant ‘cave-related habitat’ – the ‘cave threshold’ – simply because it is familiar and accessible to cavers, while ignoring all other lit, cave-related habitats.
In the world of dark holes, the physical dimensions of a potential habitat are of obvious importance in determining what creatures can colonize it. One has only to consider the relative body-diameters of a man (say 450 mm across the shoulders), a Greater Horseshoe Bat (60 mm), a cave spider such as Meta menardi (6 mm), a springtail (0.6 mm) or a nematode worm (0.06 mm) to appreciate that one creature’s spacious accommodation may be another’s unenterable squeeze, and that the cave biologist may be excluded physically from all but a tiny proportion of the very largest of cave habitats. Frank Howarth, an entomologist who works mainly on the fauna of Hawaiian lava caves, distinguishes three principal hole-size categories which appear to have biological significance for subterranean biotas. He terms these ‘macrocavernous’ (>200 mm diameter), ‘mesocavernous’ (1–200 mm diameter) and ‘microcavernous’ (<1mm diameter).
The characteristic inhabitants of Howarth’s microcaverns are sometimes termed ‘the interstitial fauna’. They include a distinctive suite of specialized, skinny-bodied crustacea (such as Bathynella and various harpacticoid copepods) and other tiny creatures (such as rotifers, nematodes and tardigrades) which mostly like to be in contact with a solid surface on all sides and typically inhabit the spaces in between unconsolidated, fine-grained sediments such as the sand and gravel of river beds and the seashore. I propose, on purely arbitrary grounds, to exclude this fauna from further discussion in this book (except for species which also frequently inhabit larger spaces), and to restrict the definition of the cave habitat to holes of 1 mm diameter upwards, that is, to mesocavernous and macrocavernous habitats.
Various vertebrates use macrocavernous caves (and the larger mesocaverns) for shelter and they, and the other species which depend on their presence, form characteristic communities which reach astonishing levels of diversity and abundance in tropical regions. I shall long remember my first visit to the spectacular Deer Cave in the Gunung Mulu National Park in Sarawak, where at dusk close on half a million bats stream out of the cave in a seemingly-endless cloud which winds its way across the sky with a rush of wings like the sound of Niagara Falls. British bat-watchers have to be content with the odd flap, but in spite of declining populations, cave-roosting bats are still widespread and bat caves do support their own suite of associated ‘batellite’ cavernicoles. Other cavernicoles may, for example, be specifically associated with the guano of cave-roosting crickets, or with cave sediments introduced by sinking streams.
Mesocavern-sized holes not only occur within karstic rocks, but also in screes, in the coarse gravels and rocky beds of upland rivers, between the pebbles and cobbles of exposed sea-shores, in the fractured zone of non-karstic rocks (especially shales) just beneath the soil, and as cooling cracks in lava flows and other igneous rocks. They represent a very much larger habitable subterranean space than do macrocaverns and so have developed a richer and often more specialized fauna, frequently dominated by species peculiar to this habitat and characterized by a reduction in the size of the eyes, loss of pigment and various other specializations. These ‘mesocavernicoles’ may also occur in soil spaces, or animal burrows, or even in large macrocaverns, provided there is an adequate food supply of down-washed organic material and a fairly stable humid microclimate. Not all species within the mesocavernous fauna will be found in all related habitats; some do not seem to occur in soil-spaces, others shun human-sized caves.
Simply as a consequence of our own species’ enormous body-size, we are physically excluded from the very habitats which are most likely to harbour a specialized fauna. In the absence of appropriate tools with which to peer inside mesocavernous habitats, cave biologists have so far been forced to infer what they can about them from the behaviour of their biotas where they pop up in the accessible portions of people-sized caves. These act as windows into the mesocavernous world, but it seems likely that they provide a distorted view, encouraging widely differing interpretations of the nature of what has been observed. The present situation in cave biology is a bit like that which prevailed among astronomers a century or so ago, when dependence on inadequate earth-based optical telescopes sustained the widely-held belief that Mars was criss-crossed by an elaborate network of irrigation canals built by Martians. Speculation and controversy abound no less in cave biology literature, while cavernicolous communities remain enigmatic and under-recorded. As a result, new species await discovery in most subterranean habitats in every part of the world including the British Isles. In short the whole subject of cave biology is very much still in its infancy. A nice illustration of this turned up on my desk in the form of a report from Frank Howarth, announcing his discovery of a brand-new diverse fauna of highly specialized cavernicoles in lava caves in tropical Australia. For years Australia was thought to have a very poor fauna of specialized cavernicoles, and a number of papers sought to explain this on theoretical grounds. For example, it was argued that Australia’s climate during the Pleistocene had not been harsh enough to exterminate the above-ground populations of its cavernicoles, and so any tendency on their part towards specialization for underground life would be continually cancelled out by gene flow from outside the cave. Having wickedly sub-titled his paper Why there are so many troglobites [= highly specialized cavernicoles] in Australia, Howarth makes the telling point that “One has to actually enter a cave and look for troglobites before proclaiming on theoretical grounds that none could exist.” I offer this creed to the reader in the context of the British cave fauna. Let us, as naturalists, devise ways to find out what lives in our underground world and get down and study it at first hand.
I have distinguished between ‘interstitial’ (microcavernous) and ‘cave’ (meso- and macrocavernous) habitats on the grounds that their biotas are substantially distinct. We might expect a similar distinction to exist between ‘aquatic’ and ‘terrestrial’ cave communities. Certainly, there are some cavernicoles which are essentially aquatic, and others which are essentially terrestrial. However, the atmospheres of most mesocavernous, and of some macrocavernous, gas-filled habitats are permanently saturated with water vapour. This poses physiological problems for many groups of terrestrial arthropods which, unless equipped to eliminate excess water from their tissues (as aquatic species do), would quickly die of ‘water poisoning’ through dilution of their body fluids. Not surprisingly, ‘terrestrial’ mesocavernicoles have been found to be physiologically specialized to cope with a hydrating atmosphere and seem able to withstand long periods of immersion in freshwater – an adaptation which is essential in habitats which are frequently flooded by downward-percolating rainwater or by fluctuations in the water-table. Some seem equally at home in air or water, and can frequently be seen feeding on the floor of cave pools among their aquatic counterparts. Conversely, many freshwater aquatic mesocavernicoles seem able to cope with ‘terrestrial’ life without undue physiological stress and have been recorded as living out of water for several weeks at a time. So we see that the distinction between terrestrial and freshwater aquatic cave habitats is not exactly cut-and-dried, although there is a clear distinction between the communities present in either zone and those found in marine cave habitats.
Not all terrestrial cave habitats are moist. Large caves with more than one entrance often experience drying airflows which can produce desert-like conditions which are lethal to the hygrophilic denizens of the mesocaverns. However, such caves are often easily accessible and attractive to vertebrates, and may (especially in the tropics) support vast populations of bats, birds and guano-associated invertebrates. Guanobious animals exhibit few or none of the morphological characteristics considered by European cave biologists to be the mark of a ‘true cavernicole’ or ‘troglobite’, yet they may be just as exclusively cave-dwelling as any mesocavern specialist. Above-ground human structures are usually designed to be as dry as possible and are seldom completely dark, and this makes them suitable as a habitat for only a very few cave-threshold specialists, such as the daddy long-legs spider Pholcus phalangioides which presumably originated somewhere in the Mediterranean region, but in the UK is found only in houses. In between the dry, draughty macrocaverns and the soggy, airless mesocaverns, there may be wide expanses of transitional cave habitats with a variable microclimate, posing a distinct set of problems for the communities which inhabit them. Terrestrial inhabitants of such places must cope with the physiological stress of desiccation some of the time and physiological drowning for the rest of the time. In the tropics, transitional cave habitats may be particularly extensive, with their own specialized faunas, often dominated by ‘bandits’ – marauding predators and scavengers which live off the scraps of the guano-based community. Climatically similar conditions are found in man-made culverts and other artificial tunnels, and these frequently attract transitional-zone cavernicoles such as the widely distributed cave spider Meta menardi. Later we shall distinguish a range of natural and artificial cave habitats principally on the basis of their microclimatic regimes.
In earlier discussing the criteria which may be important to cavernicoles in choosing their habitats, I included the apparently pedantic phrase ‘gas mix’, rather than ‘air’ in my list of the media which may fill mesocavernous voids. I did so because it seems that the atmosphere of mesocaverns may differ substantially from that found in open macrocaverns with a good air circulation (which generally have much the same atmosphere as the outside world). Bacterial decomposition of organic material in small spaces frequently results in unusually high atmospheric concentrations of carbon dioxide. Frank Howarth’s new Australian cavernicoles, mentioned earlier, were found in poorly ventilated lava caves which are thought to share the atmosphere of the mesocavernous spaces in the surrounding basalt. The air in these caves is saturated with water vapour and contains around 250 times more carbon dioxide than normal air. Bad air caves occur in Britain too, but have not yet been biologically investigated.
Finally, we may seek to distinguish cave habitats from non-cave habitats in terms of their food supply. Early cave biologists, whose experience of cave faunas was mainly confined to the larger, more easily explored ‘fossil’ macrocaverns (those no longer bearing the watercourses which formed them) of temperate European limestone areas, concluded that cave animals were perpetually starved. While food resources may be very thinly distributed in such cave habitats, in others (and particularly in the tropics) food may be superabundant. The biotas of food-poor caves are adapted to eke out what little energy is available, while those of food-rich caves are adapted to a life of plenty. Caves may contain a wide range of food sources, including living vegetation (tree roots, saprophytic plants and fungi which get their energy by digesting organic matter rather than by trapping sunlight, fruits carried in by vertebrates), living invertebrate or vertebrate animals, and all kinds of detritus. Cavernicoles may be plant-, fungus-, detritus- or bacteria-feeders, predators, parasites or a combination of these. In short, caves are more ecologically diverse than most biologists realize.
To summarize then, cave habitats may be defined as ‘perpetually-dark voids, more than one millimetre in diameter (and sometimes much larger), bounded by rock or similar inorganic materials, and filled with gas (‘fresh’ or ‘bad’ air) and fresh or salt water.’ Within such habitats, the microclimatic regime and the type and quantity of the available food-supply largely determines the species composition of the cave community. Only the largest (and often, in our islands, the least populated) cave habitats are accessible to human observers, so that we know a good deal less about the composition and functioning of cave communities in Britain and Ireland than we do about most other natural communities of our islands.
What lives in caves?
Of the voluminous literature dealing with the biota of caves, two works of this century stand out for sheer scope of vision. The first, B. Wolf’s Animalium Cavernarum Catalogus, published in three parts between 1934 and 1938, lists all animal species recorded from caves to that date. The second, by A. Vandel, published in French in 1964, discusses the biota and biology of caves worldwide. An English translation, published by Pergamon Press in 1965 as Biospeleology: The Biology of Cavernicolous Animals, is perhaps still the most useful general text despite its wacky view of evolution in caves. L. Botosaneanu’s book Stygofauna Mundi, published in 1986, gives a more up-to-date account of the fauna of subterranean waters, but there is need for a similar treatment of the terrestrial cave fauna to take into account the spate of biological discoveries in tropical caves during the decade and a half since Vandel’s book. The following brief summary illustrates the range of life forms presently known to inhabit caves.
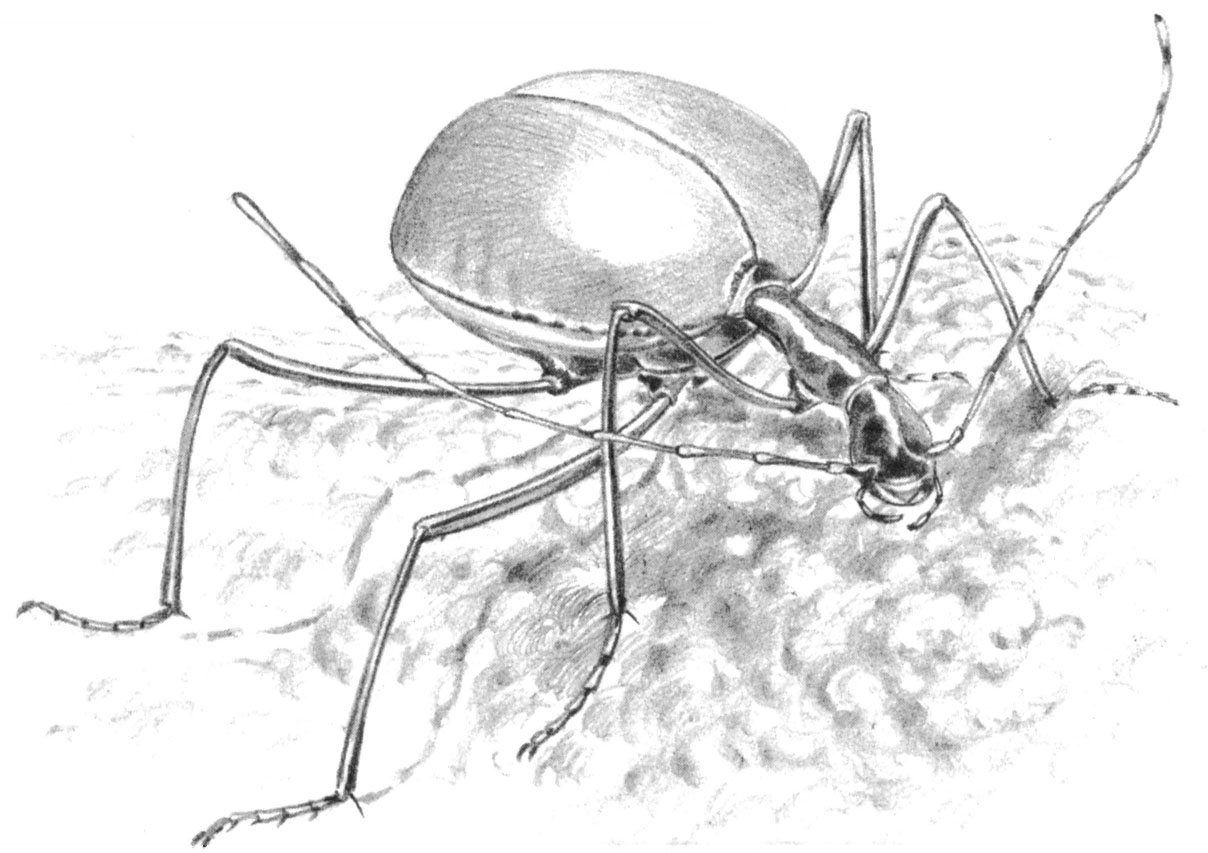
Fig. 2.2 Leptodirus hohenwarti, a highly cave-evolved beetle discovered in 1832 by the Count von Hohenwart in the Slovenian cave of Postojna Jama.
Kingdom MONERA
Phylum Bacteria
Well represented in caves. Includes saprophytes, pathogens and chemoautotrophs (which live by oxidizing or reducing iron and sulphur compounds). Bacteria are at the base of many cave food-chains.
Phylum Cyanobacteria
Some species are capable of synthesizing their pigments in the absence of light. Various Chroococcaceae are implemented in the formation of complex cave mineral deposits such as moon milk and tufa (see glossary).
Kingdom PROTISTA
Phyla Phytoflagellata, Zooflagellata, Sarcodina, Ciliophora, Sporozoa
Protista are often abundant in interstitial waters and many species occur in caves. In Turkmenistan, brackish wells in the Kara-Kum desert contain abundant populations of at least 10 species of unusually tiny, thin-shelled Foraminifera. Cave clays often contain Mastigophora, Sarcodina, Amoebina and some Ciliata.
Kingdom PLANTAE
Phylum Chlorophyta
Various free-living algae, such as Chlorella, Scenedesmus and Pleurococcus are found growing deep inside caves. Though able to synthesize pigments in the dark, they appear to use non-photosynthetic metabolic pathways. The other plant phyla Rhodophyta, Phaeophyta, Bryophyta are essentially absent from the dark parts of caves. The phylum Tracheophyta is represented by a very few aberrant saprophytic species which can live independently of sunlight.
Kingdom FUNGI
Phyla Zygomycetes, Ascomycetes, Basidiomycetes, Myxomycetes
Fungi are important in cave ecosystems. Most are saprophytic on organic material washed into caves and form the main food base of cave communities. Some are epizoic (live on the outer surface of animals) or parasitic. Most members of the phylum Oomycetes are parasitic on flowering plants and therefore not represented in caves.
Kingdom ANIMALIA
Phylum Porifera
Encrusting sponges may be the commonest organisms in tidally flushed submarine caves.
Phylum Coelenterata
Hydra viridissima occurs in groundwaters in the Southern Carpathians of Europe. Marine Hydrozoa and Anthozoa are commonly found in submarine caves.
Phylum Platyhelminthes
Rhabdocoel Turbellaria are common in wells, springs and groundwaters. Cave-evolved Triclads have a worldwide distribution, with most species within three planarian families: Dendrocoelidae, Kenkiidae and Planariidae.
Phylum Nematoda
Free-living nematodes are frequent in groundwaters, caves and mines worldwide. Several freshwater species of the otherwise exclusively marine Desmoscolecidae inhabit caves in Slovenia.
Phyla Nemertinea and Rotifera
A few species of these small creatures inhabit interstitial waters and caves.
Phylum Annelida
Submarine caves often contain huge populations of sedentary polychaete worms and a number of cave-evolved freshwater polychaetes are known from Switzerland, Slovenia, Japan and Papua New Guinea. Oligochaete worms are often abundant in caves, in groundwaters and sediments. The family Lumbriculidae contains many essentially cavernicolous species. Cavernicolous leeches are known from Central Europe and several tropical countries.

