




Полная версия
Психотроника и днк. Искусственный интеллект в битве со старением организма человека. Психотроника и днк
Специфика расположения уровней допускает их инверсную заселенность, что позволяет использовать углекислый газ в качестве активной среды лазера (Л ~ 10 мкм). В жидких средах предрасположенных к самоорганизации молекула С02 может быть донором колебательных квазифотонов для молекул с карбоксильной группой (-НСО). Аналогично, молекулы с ароматическими циклами будут акцепторами квазифотонов, отвечающих конформационным колебаниям изоморфных им насыщенных углеродных циклов и гетероциклов. Высокая активность, например, стероидных гормонов производных холестерола, имеющих конденсированные гексановые цикла, может быть обусловлена насыщенностью их молекул квазифотонами с зр3-метрикой (рисунок тринадцать). При этом изоморфные фрагментам гормонов ароматические молекулы (бензол, антрацен, пирен), эффективно дезактивируя гормоны и искажая их метаболические функции, могут инициировать канцерогенез.
Метаболические квазифотоны
Для унификации языка биоэнергетики соотнесем с величинами энергий химических связей, колебательно-вращательных и тепловых движений атомов и молекул энергию квазифотонов соответствующей метрики. Дееспособность мозга обеспечивает энергия ферментативные реакции окисления глюкозы в митохондриях и анаэробного ее гликолиза в глазном яблоке. В этих реакциях электромагнитная – энергия химических связей глюкозы и кислорода трансформируется в энергию макроэргических связей АТФ, которая в последующих реакциях гидролиза АТФ преобразуются в кинетическую и колебательно-вращательную энергию метаболитов и молекул среды. Химическая активность этих молекул реализуется затем через действия их энергии возбуждения, которую и моделируют квазифотоны соответствующей энергии и метрики.
Суммарный энергетический эффект всех стадий ферментативной реакции окисления глюкозы в митохондриях имеет своим пределом тепловой эффект реакции горения глюкозы в атмосфере кислорода:
C6Hi206 +6 02 -> 6 С02 +6 Н20 +2800 (кДж/моль). (12)
Реакция окисления глюкозы в митохондриях сопряжена с реакцией синтеза АТФ, при этом на одну молекулу глюкозы приходится 38 тридцать восемь молекул АТФ. При анаэробном гликолизе глюкозы образуются только две молекулы АТФ и две молекулы хиральной молочной кислоты, которые, очевидно, вносят свой вклад в хиральность энергетики глаз и мозга. Максимальный выход метаболической энергии даст гидролиз 38 тридцать восемь молекул АТФ по схемам:
АТФ -> АДФ + Р~Р +36 (кДж/моль)
Р~Р -> Р + Р +33,4 (кДж/моль).
Полная энергия макроэргических связей 38 тридцати восьми молекул АТФ равна 2640 кДж/моль, что составляет ~95% девяносто пять процентов от предельного значения энергии сгорания одной молекулы глюкозы. Это говорит о высокой эффективности ферментативных реакций трансформации квазифотонов, соответствующих о-связям С-С, С-О-С, С-Н глюкозы в квазифотоны, локализованные на двух макроэргических связях Р~0- в АТФ.
Предположим, что квазифотоны равновероятно распределяются по связям продуктов реакций окисления глюкозы и гидролиза АТФ, тогда предельные значения энергий квазифотонов, отвечающих данным реакциям будут равны 1/12 и 1/152 от теплового эффекта реакции (12), равного 4,5 10~18 Дж, то есть ~3 10~19 и ~3 Ю-20 Дж, соответственно. Если к этим квазифотонам применить универсальное соотношение между энергией и характерным размером (г) дискретного элемента материи (v/g-napa, элементарная частица):
Е ~ fic/r, (13)
то для квазифотона, действующего в виде кванта метаболической энергии, получим радиус ~1 мкм, сравнимый с радиусом аксона.
Метаболические квазифотоны могут принимать активное участие в ферментативном синтезе белков и нуклеиновых кислот, а также в репликации и транскрипции ДНК. Можно представить участие квазифотонов в расплетении двойной спирали ДНК следующим образом. В области репликативной вилки сахарофосфатный остов цепи ДНК резонансно поглощает метаболические квазифотоны колебательного типа. Возрастает упругость цепей, что и приводит к разрыву водородных связей между ними. Учитывая, что на два сахарофосфатных звена спирали ДНК приходится одна водородная связь и ее энергия равна ~19 кДж/моль (3 Ю-20 Дж), получится, что для ее разрыва достаточно поглощения цепью ДНК одного метаболического квазифотона.
Присутствие изоморфных аминокислотных фрагментов в пептидных цепях белка и в структуре нейромедиаторов (глицин, ацетилхолин, глутаминовая кислота, дофамин, серотонин и др.) позволяет предложить резонансный механизм передачи квазифотона колебательного типа при контакте нейромедиатора с рецептором. Из-за наличия в структурах медиаторов электроно-, протонодонорных и акцепторных групп их основное электронное состояние характеризуется внутримолекулярным переносом заряда Д+б-С-А'6. Здесь Д- аминогруппы, метоксигруппа, бензольное кольцо и А – карбонильная и гидроксильные группы, а С – цепочка из о-связей. Этот фактор и предрасположенность медиаторов к образованию водородных связей лежат в основе их физической и химической сорбции на рецепторах постсинаптических мембран. Рецептор, принимая или отдавая квазифотон при контакте с нейромедиатором, меняет свою конформацию, запирая или открывая при этом кальциевый канал мембраны.
Термодинамика мозга
Мозг в целом можно считать реакционной термодинамической системой, находящейся в стационарном состоянии. Приток энергии и сброс избыточного тепла мозгом сбалансированы в узком диапазоне температур от~37°(центр мозга) до ~36°С (кора мозга). Этот градиент температуры, будучи обусловлен более низкой температурой внешней среды, может играть существенную роль в ориентировании тепловых потоков внутри мозга. Аналогичный градиент температуры наблюдается и для тела, она имеет максимум в прямой кишке, а минимум в поверхностном слое клетчатки и мужских яичках. Диапазон оптимальной температуры метаболизма находится в пределах значений температур, для которых изобарная теплоемкость чистой воды имеет минимум. Особенности термодинамики фазовых переходов водных растворов в процессе филогенеза легли в основу механизма адаптации живых систем, которая, по сути, представляет собой изоэнергетические переходы или переходы с энергией активации порядка кДТ (при АТ ~ ОД – 1 К) между состояниями разной степени упорядоченности белковых молекул или однородных, молекулярно-клеточных ансамблей. Снижение энтропийной составляющей внутренней энергии живой системы сопряжено с резонансным поглощением ею кванта внешней электромагнитной или нейтринной энергии, который она преобразует в активный метаболический квазифотон. Жидкостная среда обеспечивает отвод кванта тепловой энергии (энтропии) за границы системы, а действием квазифотона реализуется функция той или иной структуры мозга, включающей в себя упорядоченную подсистему.
Энергия, выделяемая или поглощаемая при таких переходах, может оказаться намного порядков меньше кТ. В неравновесных условиях колебания отдельных макромолекул могут синхронизироваться, в частности, посредством электромагнитного поля.
Таким образом, термодинамика мозга сочетает равновесно-стационарную термодинамику метаболизма и неравновесную термодинамику нейросети, «рабочего телом» которой является Бозе-газ квазифотонов. Соответственно, внутренняя энергия U мозга как функция его состояния будет зависеть в общем случае от температуры (или энтропии S), от тензора деформаций G, зависящего от внутричерепного давления, от магнитного момента М отдельных метаболитов и макроструктур, от суммарного момента количества движения ядер и атомов L и от поляризация среды Р. Следовательно, полный дифференциал внутренней энергии U- U (S, G, М, L, Р) будет иметь вид:
dU= TdS – ndG + BdM + DdP + FdJ, (14)
где T – абсолютная температура системы; П – тензор давлений; F – вектор ориентационной поляризации системы спинов или моментов импульса. В выражении (14) член FdL характеризует работу, связанную с ориентационной поляризацией системы ядерных спинов или моментов импульса атомов и молекул (подобно тому, как члены DdP и BdM определяют работу, связанную с поляризацией и намагничиванием системы).
Ориентационные и поляризационные эффекты существенную роль играют в инициации фазовых переходов в однородных газовых и жидкостных системах мозга и организма. Высокую чувствительность данных систем к параметрам входящим в (14) обеспечивает хиральность метаболитов (в основном сахаров) и физико-химические особенности молекулярной и жидкой воды. Такие системы формируются в следующих структурах и средах организма и мозга:
– желудок, матка, трахея, черепно-лицевые пазухи, полость эпифиза; -оболочки и желудочки мозга, венозные синусы, глазное яблоко;
кровеносная и лимфатические системы;
паренхима органов (легкие, печень, селезенка, яички, женская грудь);
подкожная клетчатка, соединительная и костная ткань.
Все перечисленные системы в норме функционируют в двух режимах- стационарном (квазиравновесном) и неравновесном. Первый характерен для бодрствующего состояния организма и мозга не занятого мыслительной работой, а второй режим соответствует состоянию сна или творческой работе. В первом режиме обмен энергией со средой происходит непрерывно, а во втором – квантуется. Механизм акцепции кванта внешней энергии в фазовом переходе кооперативной системы иллюстрируют процессы конденсации паров воды в точке росы и квантовой Бозе- конденсации.
Акцептированию квантов энергии микроволнового – диапазона или нейтринной энергии в указанных средах способствует снижение температуры организма во сне на ~1К, а также пониженные температуры стекловидного тела глаз, периферийной (депонированной) крови и яичек. В акцепции хиральных квантов нейтринной энергии большую роль играют сахара, содержание которых в крови возрастает в утренние часы до восхода солнца. В это время нейтринная составляющая солнечного излучения отфильтровывается от электромагнитного – излучения поверхностным сегментом коры земного шара. Важную роль в акцепции энергии стекловидным глазом играет полисахарид – гиалуроновая кислота. Почти половина всей гиалуроновой кислоты организма человека сосредоточено в его коже, где она располагается в соединительной ткани дермы между волокнами коллагена и эластина, а также в клетках рогового слоя корнеоцитах. В дерме содержится 70% воды, что составляет ~20% двадцать процентов всей воды организма. Поглощаемая организмом энергия, конденсируясь на метаболитах в составе жидких сред (кровь, спинномозговая жидкость), передается в мозг по нейрогуморальным и воздушным каналам (из легких).
Поглощение внешнего электромагнитного – кванта и формирование квазифотона из энергоформ в общем случае подчиняется фрактальнорезонансному механизму и принципу изоэнергетичности. С учетом (13) принцип изоэнергетичности для резонансных взаимодействий и фазовых переходов в кооперативных системах можно выразить соотношением:
hC/r — N (TiC/R), (15)
здесь г характеризует метрику квазифотона, a R – энергоформы и г = R/N; число N принимает любые значения меньшие числа Авогадро при конденсации энергоформ в квазифотоны и достигает числа Авогадро при участии энергоформ в слабых взаимодействиях.
То обстоятельство, что между тепловой формой движения и ориентацией спинов существует определенная связь, еще не дает оснований приписывать эту форму спиновой системе, тем более что охлаждение конденсированных сред до температур, близких к абсолютному нулю не приводит к исчезновению собственного момента вращения ядер. Это обстоятельство также свидетельствует о недопустимости описания спиновой системы параметрами термической степени свободы и о расхождении такого описания со вторым началом термодинамики для квазистатических процессов (принципом существования энтропии).
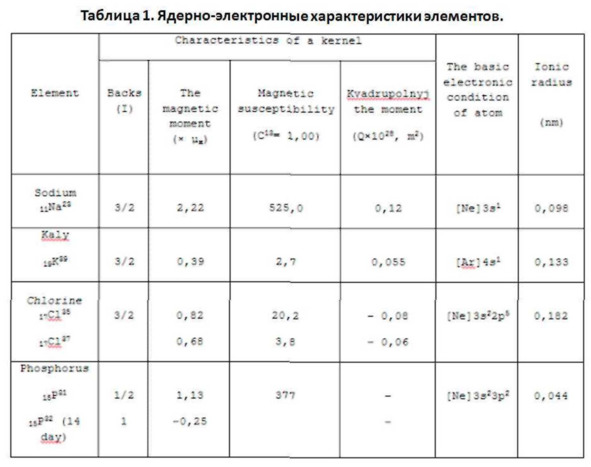
Еще одним подтверждением несводимости спин-спинового взаимодействия к теплообмену являются, как ни странно, те самые опыты по «смешению» двух систем противоположно ориентированных спиновых систем (7Li и 19F) кристалла LiF Эти опыты показали, что «температура» смеси отнюдь не подчиняется обычным для таких случаев законам сохранения вида:
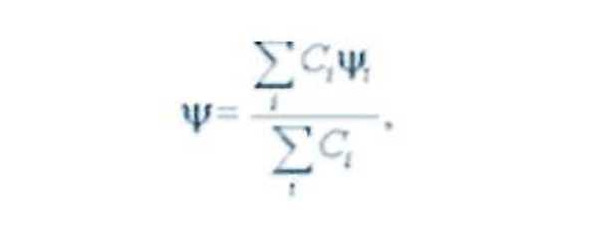
где ф, – какой-либо интенсивный параметр (температура, химический, электрический, гравитационный и др. потенциал); С, – соответствующий экстенсивный параметр (полная теплоемкость, число молей, заряд, масса и т.п.). Напротив, в случае спиновой системы в выражении со «спиновой теплоемкостью» С, сопряженная величина, обратная абсолютной температуре. Отсюда следует, что законам типа этой формулы подчиняется не температура, а ядерная намагниченность М, относящаяся к иной степени свободы спиновой системы. Вместе с тем было бы также ошибочным сводить спин-спиновое взаимодействие к торсионному (порожденному различной плотностью углового момента вращения).
Такое взаимодействие определенно имеет место в вязких средах, обладающих некоторым моментом инерции (как, например, в гидромуфтах). Однако наличие такого взаимодействия в вакууме пока экспериментально не доказано. Кроме того, в отличие от торсионного спин- спиновое взаимодействие проявляется и в тех случаях, когда вращающиеся объекты обладают одинаковой плотностью угловых моментов вращения (в частности, одинаковой угловой скоростью), поскольку гироскопический эффект проявляется и в этом случае. Поэтому его следует отнести (наряду с электромагнитным, гравитационным, сильным и слабым взаимодействием) к еще одному независимому виду взаимодействия. Способность его передавать упорядоченность одних микрочастиц другим, а также сравнительно большие времена спин-спиновой релаксации могут пролить новый свет на ряд не познанных до сих пор явлений. К ним относятся процессы воспроизводства или изменения структуры объектов живой и неживой природы, эффекты «памяти» воды (в том числе появление у нее лекарственных свойств при «перезаписи» на нее структуры этих лекарств), лечебный эффект приборов, генерирующих различные поля, или геометрических фигур, изменяющих диаграмму направленности разнообразных излучений, многочисленные проявления «фантомов» (призраков) отсутствующих тел и т. п. Однако рассмотрение этих вопросов выходит далеко за рамки темы.
Организации мозга
Функциональная иерархия мозга
Функциональная иерархия мозга человека строится на физических свойствах следующих его структур: неокортекс, базальные ядра, лимбический мозг, таламус, гипоталамус, гипофиз, эпифиз, ретикулярная формация и мозжечок. К отдельным элементам иерархии следует отнести жидкостные системы мозга (кровеносная и ликворная), а также весь комплекс внутричерепных нервных коммуникаций, выделив в нем мозолистое тело, зрительный нерв и лучистости таламуса.
В структуре коры различают поверхностные специализированные зоны и шесть слоев. Самый верхний слой образуют горизонтально ориентированные апикальные дендриты пирамидных клеток и аксоны звездчатых клеток, которые обеспечивают внутрикорковые связи между соседними нейронами. Горизонтальной ориентации диполей нейронов данного слоя соответствует динамичное электрическое поле, которое может играть роль защитного электромагнитного – экрана. Остальные пять горизонтов коры структурируются сначала в нейро-глиальные модули (диаметр ~ 100 – 150 мкм), а затем, в колонки диаметром до 1 мм и со средним числом нейронов ~100 шт. Синхронизация электрической активности нейронов в модулях и колонках приводит к формированию в пучках отходящих от них аксонов залповых импульсов. Процессу суммирования потенциала действия в залпы может предшествовать конденсация по (15) внешних электромагнитных – квантов или квазифотонов стимулов потенциала действия на отдельных нейронах в модулях, а затем в колонках. Например, при конденсации ~100 квазифотонов с характерным радиусом 100 мкм в залповом импульсе может образоваться квазифотон радиуса 1 мкм (энергия ~10~19 Дж).
ЗАВИСИМОСТЬ ОТ ТЕМПЕРАТУРЫ ОПТИЧЕСКОЙ АКТИВНОСТИ ФИЗИОЛОГИЧЕСКИХ РАСТВОРОВ САХАРОВ
Исследована зависимость от температуры оптической активности водных растворов метаболических сахаров и определена энергия активации реакции их ассоциирования в надмолекулярные хиральные структуры. Ее величина оказалась близка к значениям энергии вращательного движения и биогенного микро волнового-излучения. Исходя из этого и учитывая зависимости оптической активности сахаров от времени и даты, места и ориентации прибора, приняли за основу механизма действия хирального фактора фило- и онтогенеза метаболический эффект реакции образования хиральных водно-сахарных ассоциатов. Оптически активные D-caxapa являются основным источником энергии метаболизма живых систем. Обязательное посредничество ахиральной молекулы аденозинтрифосфата (АТФ) в процессе метаболизации энергии химических связей сахаров нивелирует их хиральность, унифицируя тем самым энергию макроэргических связей АТФ. Однако роль сахаров в возникновении и развитии жизни не ограничилась их вкладом в биоэнергетику. В ходе эволюции усложнялись физико-химические свойства сахаров и, соответственно, возрастала их роль в энергоинформационном обмене живых систем с внешней средой. Моно- и полисахариды вошли в состав структурных элементов биомолекул, клеток, жидких сред, информационных систем, они регулируют иммунитет высших организмов. Учитывая морфо – функциональную универсальность D-сахаров, можно предположить, что именно их физико-химические свойства обеспечивают высокую чувствительность механизма адаптации живых систем к внешнему постоянно действующему хиральному фактору.
Следует отметить, что вопрос о физической природе хираяьного фактора и механизме его влияния на биогенез до сих пор остается открытым. Впервые данную проблему сформулировал Пастер в девятнадцатом веке, предположив, что дисимметрия живых систем возникла на ранних стадиях биологической эволюции под действием электрических и магнитных полей космического происхождения. В обоснование этой идеи в работе приведен возможный механизм асимметричного действия электромагнитного поля на макромолекулу белка. Согласуется с нею и гипотеза о ключевой роли сахаров еще на пребиотическом этапе биогенеза. Привлекательна также идея о нейтринной природе хирального фактора, которую можно соотнести с гипотезой Вернадского о детерминировании дисимметрии живых систем хиральностью физического вакуума (эфира). Реликтовое нейтрино, составляя энергетическую основу вакуума, вполне может обеспечивать его гравитационную и электромагнитную динамику (эффект Фарадея, например).
В общем случае в основе механизма чувствительности живой системы к хиральным факторам помимо хиральности ее элементов должно лежать то или иное взаимодействие между ними, обеспечивающее переход системы в коррелированное состояние с новым качеством. Переход в данное состояние (фазу) ведет к возрастанию порядка системы или снижению энтропии и при этом увеличивается ее устойчивость (время жизни). За перестройку порядка системы на микро и макро уровне ответственны электромагнитные взаимодействия между элементами. Динамику, кинетику, радиус действия и стереометрию упорядочивающих сил в живых системах лимитируют молекулярно-кооперативные свойства воды.
С целью получения дополнительной информации о вкладе сахаров в механизм чувствительности живых систем к хиральному фактору в настоящей работе исследовали температурные и концентрационные зависимости оптической активности водных и водно – этанольных растворов 0-глюкозы и сахаров. Данные растворы моделировали состав физиологических жидкостей (межклеточная жидкость, лимфа, ликвор, синовия). Исследовали оптическую активность композиции: ФР + хондроитинсульфат (10%) + гиалуроновая кислота (0,8%) + сахар (10%) {ФР + ХС (10%) + ГК (0,8%) + сахар (10%)}, которая моделировала высокомолекулярное соединение, играющее активную роль в энергетике соединительной ткани. Величина U раствора ФР + ХС (10%) при комнатной температуре (Тком) составила -2,70°(кювета 100 мм). Пищевой сахар ([a] D = 66,5°) растворяли в физрастворе. Полученные результаты, согласуясь с известными данными, свидетельствуют о чувствительности оптической активности растворов к положению Солнца и, по-видимому, к фазе Луны. Для количественной обработки результатов предположили, что в растворах сахаров при достижении порогового значения концентрации, наряду с оптической активностью отдельных молекул свой вклад в хиральность раствора вносят и надмолекулярные их образования. В пределе межмолекулярных взаимодействий характерных для кристалла сахара его удельное вращение приобретает анизотропию (1,6 – 5,4 град/мм для различных направлений) и почти на порядок превышает удельное вращение растворов сахара. О возможности вклада межмолекулярных взаимодействий в хиральность раствора говорит еще такой факт. Высокое значение удельного вращения кристалла кварца (21,7 град/мм) обусловлено только дальним порядком его спиральных полимерных цепочек, мономерами которых являются ахиральные тетраэдры из молекул Si02.
Очевидно, что в случае растворов сахаров энергия межмолекулярных взаимодействий (Е) будет определяться средним расстоянием между молекулами, то есть их концентрацией, а также их ориентацией относительно друг друга и геометрией надмолекулярных образований воды. При концентрации сахаров в растворах 20 – 40% (~2 моль/л) среднее расстояние между молекулами, равное ~10—7 см, сравнимо с их размерами. В этих условиях сахара должны ассоциировать, образуя упорядоченные надмолекулярные структуры под влиянием сил Ван-дер-Ваальса и при посредничестве динамичных водородных связей между молекулами воды. Процесс ассоциирования будет сочетать в себе вращательную корреляцию самих сахаров с пространственной корреляцией включенных вассоциат динамичных кластеров воды. Причем на обеих корреляциях будет сказываться хиральность электронной структуры молекулы сахара, в результате чего ассоциат и приобретает собственную хиральность, увеличивая тем самым суммарную оптическую активность раствора. Таким образом, влияние температуры на динамику ассоциирования сахаров может обусловить температурную зависимость а их высококонцентрированных растворов. Суммарный эффект кооперации сахаров и молекул воды будет максимален при самой низкой температуре опыта, а значит, максимальным будет и значение а.
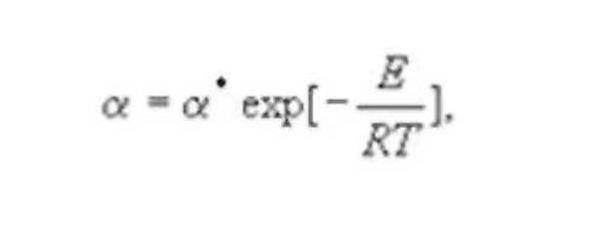
где Е-энергия взаимодействия или корреляции молекул. Наличие излома на некоторых температурных зависимостях при ~295К может быть связано с изломом в этой критической точке температурной зависимости динамической вязкости воды. Этот результат подтверждает зависимость динамики кооперативных взаимодействий растворенных молекул от степени упорядоченности самой воды, которая возрастает по мере приближения температуры раствора к критическим точкам (~22, ~36°С). О том же говорит увеличение величин Е при добавлении в растворы глюкозы и декстрана этанола, молекулы которого могут оказывать стабилизирующий эффект на водородные связи в динамичных кластерах воды. Известно, что ассоциирование стимулирует добавка NaCI и в согласии с этим величина Е для физраствора сахара больше, чем Е для его водного раствора. Был зафиксирован локальный скачок величины раствора ФР + ГК (0.8%) + сахар (20%) при неизменном значении, который явно не коррелировал с изменениями горизонтальных составляющих геомагнитного поля.
Упорядочивающий эффект фазового перехода вхиральной живой системе, очевидно, сопряжен с резонансной синхронизацией крутильных колебаний молекул и последующей конденсацией квантов энергии порядка Ещ, Ерез,
Емкв и Er в квант энергии порядка энергии макроэргической связи. В принципе, по такому же механизму кванты энергии порядка Е0 могут конденсировать в кванты тепловая критическая и Крутильные колебания больших молекул в момент пространственной корреляции однотипных электронных орбиталей в молекуле полисахарида, имеющей в своем основании гиалуроновую кислоту. Очевидно, что такие переходы идут в информационных системах живого организма, имеющих однородно множественную молекулярно-клеточную структуру, предрасположенную к гелеобразованию. Ew– крутильные колебания больших молекул в конденсированной фазе (w ~ 1011 с 1) / Ерез – энергия резонансного возбуждения, Емкв. биогенное микро волновое излучение – излучение, Ек. реликтовое излучение, Е0– энергия нейтринных флуктуаций вакуума. К таким системам относятся, например, стекловидное тело глаза, цитоплазма аксона, синовия. Сами же фазовые переходы в информационных системах наиболее вероятны в состоянии сна, когда снижается температура организма. По-видимому, в основе парадоксальной фазы сна, характеризуемой быстрым движением глаз (фаза – быстрого движения глаз), тоже лежат фазовые переходы, обеспечивающие корректировку гомеостаза внешними и внутренними хиральными факторами.
Результаты позволяют заключить, что предрасположенность физиологических жидкостей содержащих метаболические сахара к фазовым переходам в критических температурных точках воды, может лежать в основе механизма адаптации живых систем к изменениям внешних физических условий, как в геологическом, так и в реальном масштабе времени. Перенос информации в организмах, чаще всего, выполняется волнами разной природы, при чем и волны, и методы переноса ими информации (способы модуляции) оказываются специфичными, не такими, как в технике. Биологические информационные системы являются системами молекулярного уровня. Сюда относятся наши сенсоры – зрение, слух, обоняние и т. д. Но сюда относятся и другие информационные системы организмов – системы управления геномом, мозг, нервная система в целом. Поскольку биологические информационные системы основаны на процессах молекулярного уровня, эволюция четко привела их механизмы переноса информации к использованию медленно распространяющихся волн. Скорость распространения химических структурогенных волн в многоклеточных организмах, по расчетам, около 4 км/с. что в 75.000 раз меньше скорости электромагнитных волн в вакууме. Оболочка ядра преобразует энергию химических волн в энергию акустических колебаний, скорость распространения которых около 1.5 км/с. т.е. в 200.000 раз ниже скорости света.