


Полная версия
Парадоксы эволюции. Как наличие ресурсов и отсутствие внешних угроз приводит к самоуничтожению вида и что мы можем с этим сделать



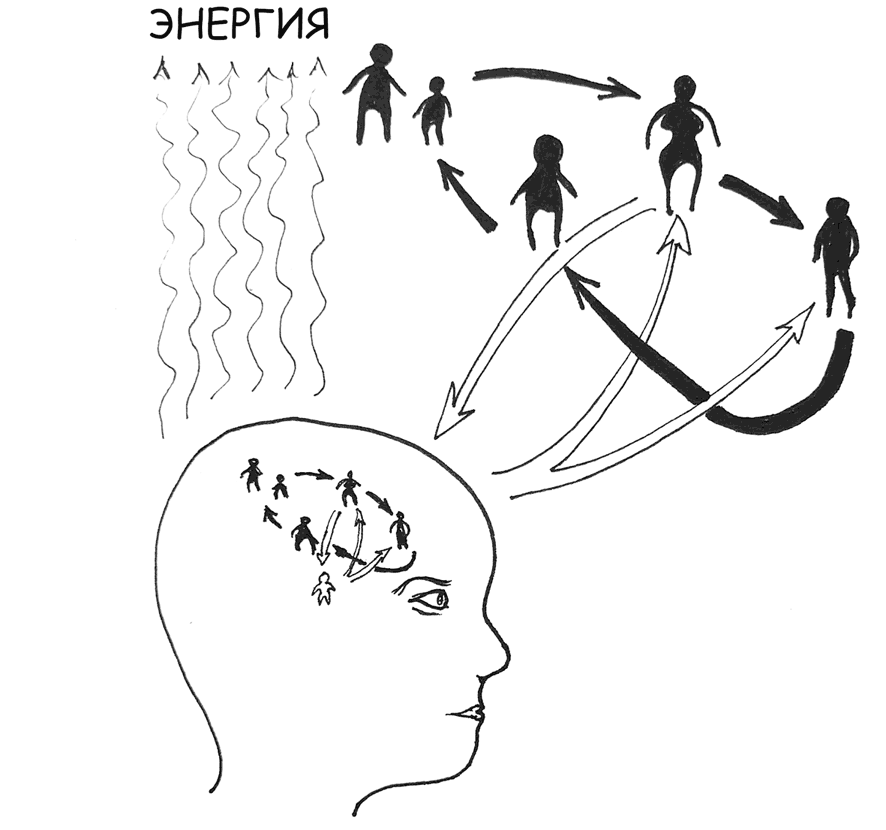
Рис. 9. Чрезмерный расход энергии при развязывании «узлов социализации» как возможная причина депрессии
То, что физические упражнения у человека сами по себе практически не влияют на снижение веса, совершенно не означает, что они не нужны; наоборот, они критически необходимы для поддержания нашего здоровья.
Понцер сравнивает необходимость движения для людей с пассивным дыханием у акул: у этих хищников в процессе эволюции выработалось крайне агрессивное пищевое поведение; охотясь, они без остановки плавают и днем, и ночью. Постоянное движение вперед обеспечивает постоянное заглатывание воды и пропускание ее через жабры, что эволюционно привело к утрате развитой жаберной мускулатуры. Это позволяет экономить энергию, но ведет к необходимости постоянного движения: остановившись, многие виды акул «задыхаются». Последствия гиподинамии для человека гораздо менее стремительны, но не менее губительны. Однако, в отличие от акул, движение еще не стало для человека комфортным состоянием, подавляющему большинству из нас приятнее находиться в состоянии умеренного покоя. По выражению палеонтолога Нила Шубина, в строении и функционировании каждого из нас можно увидеть наших эволюционных предков, в анатомии, например «внутреннюю рыбу» (Neil Shubin, 2008). В свою очередь, по мнению Германа Понцера, «внутренняя обезьяна» в нашей физиологии зовет нас в потерянный рай жизни в джунглях, где, как и наши двоюродные братья шимпанзе и гориллы, можно 8–10 часов в день тратить на отдых, груминг и еду, спать 9–10 часов, бродить взад-вперед не более 3 км и пролезать вверх-вниз не более 100 м. Но почему-то около 2 млн лет назад наши непосредственные предки-гоминины буквально выбежали из этого рая. И только около 200 лет назад человечество начало снова физически «приостанавливаться». Для застрявшей в комфорте части человечества движение снова становится жизненно необходимым хотя бы для того, чтобы не набирать вес в ожирении (и это не противоречит тому, что физические упражнения сами по себе, без сокращения диеты, плохо справляются с УЖЕ накопившимся ожирением), предотвращать кризис иммунной системы, особенно в пожилом возрасте, поддерживать когнитивную функцию, противостоять, в конце концов, сердечно-сосудистым заболеваниям – главной причиной смертности в XXI веке.
Толкаем гири, но не забываем про соседей
Однако продемонстрированное влияние физических упражнений как горметического стресса на состояние здоровья умножается на 0 или на 10 одним очень важным фактором, находящимся в настоящее время в фокусе внимания сотен исследовательских групп – состоянием микробиома, в первую очередь микробиома кишечника. Микробиом может в значительной степени модулировать физиологическую результативность физических упражнений через ряд сравнительно хорошо исследованных механизмов, связанных, например, со стимуляцией вегетативной нервной системы. Реакция на стресс, в том числе горметический, определяется статусом этой системы, находящейся под постоянным влиянием оси взаимного влияния «мозг-кишечник», где роль ведущего канала взаимодействия играет блуждающий нерв. Функциональность этой оси также «администрируется» через выработку нейротрансмиттеров, таких как гамма-аминомасляная кислота (ГАМА), нейропептид Y, серотонин, а также биологически сверхактивных короткоцепочечных жирных кислот, и оборот этих соединений в значительной степени контролируется кишечной микрофлорой (БОН: глава VIII). Фундаментально кишечная микрофлора также контролирует всасывание питательных веществ, то есть, по сути, общее поступление энергии в организм хозяина.
Менее исследованным, но более интересным, и, на мой взгляд, гораздо более многообещающим в силу возможного синергического эффекта на здоровье является обратное влияние физических упражнений на состав и состояние микрофлоры. Наиболее часто в результате систематических физических упражнений (см. обзор Clark A., Mach N., 2016) исследователи обнаруживают рост микробного разнообразия кишечника, снижение соотношения Firmicutes/Bacteroidetes (хотя есть и прямо противоположные результаты), увеличение содержания бактерий рода Akkermansia и бактерий-производителей масляной кислоты, то есть всего того, чего энтузиасты главенства кишечного микробиома в его симбиозе с человеком пытаются достичь самыми разнообразными и неординарными способами, включая пересадку кала от «здоровых» индивидуумов больным.
В качестве предполагаемых механизмов различные группы исследователей называют ускорение транзита пищи через ЖКТ (Oettlé GJ, 1991), изменение профиля желчных кислот (Hagio M. et al., 1985), увеличение выработки короткоцепочечных жирных кислот (Cerda B., 2016), модулирование сигнальных путей клеточных иммунных toll-like рецепторов (Fracaux M., 2009; Frosali S., 2015), изменение уровня защитных секреторных иммуноглобулинов А (Viloria M. et al., 2011), количества В-лимфоцитов и CD4+ Т-лимфоцитов, и даже, парадоксальным образом, через вызванную нагрузками потерю веса (Turnbaugh P. J. et al., 2006). Остается вопрос, какой из механизмов в этой сложной комбинации эффектов все-таки является ведущим. Связующим звеном взаимного влияния физических упражнений на кишечный микробиом и микробиома на эффективность упражнений может оказаться, например, обнаруженное торможение выработки белков теплового шока в клетках кишечного эпителия в результате длительных упражнений, что ведет к упрочению плотных связей между ними и снижению проницаемости кишечника (Dokladny K. et al., 2016; Mailing L. J., 2019).
Не будет удивительным, что взаимное положительное влияние микрофлоры и клеток кишечника в значительной степени опосредованно митохондриями последних, что особенно проявляется в ходе физических нагрузок (Clark A. and Mach N., 2017). Так, микрофлора снабжает митохондрии их любимыми короткоцепочечными жирными кислотами (КЦЖК) – масляной (бутиратом) и уксусной (ацетатом). Благодаря бутирату «накачанные» митохондрии активнее вырабатывают PGC-1α и АМРК, что в данном случае «успокаивает» провоспалительный настрой клетки, связанный с NF-кВ, и стимулирует биогенез митохондрий (нарастание их количества и массы). Кроме того, вместе с вторичными желчными кислотами КЦЖК позволяют выстроить в просвете кишечника оптимальный окислительно-восстановительный потенциал, необходимый как для наилучшей производительности ЭТЦ, так и самочувствия кишечных бактерий. Благодаря оптимизированной функции митохондрий, в свою очередь, улучшаются иммунные свойства слизистой кишечника, ее способность противостоять возможному вторжению патогенов. Очень важным комплексным эффектом нормально функционирующих митохондрий в клетках кишечной стенки является нормализация ее проницаемости.
Летучие мыши, чей образ жизни, как обсуждалось, в плане физической нагрузки достаточно сильно отличается от остальных млекопитающих, также весьма основательно отличаются от других млекопитающих и по типичному составу микрофлору, приближаясь в этом отношении к птицам – животным, в чем-то более к ним близким по образу жизни и характеру физической нагрузки (Lutz H. L. et al., 2019).
В итоге оказывается, что физическая нагрузка в качестве горметического стресса, действуя на весь организм в самом широком смысле, то есть включающем его микробиом, запускает процесс благотворной адаптации также всего организма в самом широком смысле, что может как кратно увеличивать, так и нивелировать эффективность нагрузки. Это простое рассуждение приводит к необходимости постараться рассмотреть вопросы энергии и движения, в том числе движения эволюции, в контексте постоянного тесного взаимодействия взаимозависимых геномов.
Библиографический список
1. Селье Г. (1982). Стресс без дистресса. – М: Прогресс.
2. ФГБУН «ФИЦ Питания, биотехнологии и безопасности пищи» (2020). Оценка влияния регулярного употребления воды с высоким содержанием бикарбоната натрия на анаэробную производительность. Отчет о научно-исследовательской работе.
3. Александер Д., Райхлен Д. (2020). Зачем мозгу физические упражнения? ВМН, № 3, с. 5–11.
4. Понцер Г. (2017). Парадокс физической активности. ВМН, № 4, с. 27–33.
5. Понцер Г. (2019). Эволюция и тренировки. ВМН, № 3, с. 5–13.
6. Шубин Н. (2008). Внутренняя рыба. История человеческого тела с древнейших времен до наших дней. – М.: Династия.
7. Radak Z., Chung H. Y., Koltai E., Taylo, A. W., Goto S. (2007). Exercise, oxidative stress and hormesis, Ageing Res. Rev. 170.
8. Malm C., Sjodin T. L., Sjoberg B., Lenkei R., Renstrom P., Lundberg I. E., Ekblom B. (2004). Leukocytes, cytokines, growth factors and hormones in human skelet al. muscle and blood after uphill or downhill running. J. Physiol. 556, 983–1000.
9. Peake J. M., Nosaka K., Muthalib M., Suzuki K. (2006). Systemic inflammatory responses to maximal versus submaximal lengthening contractions of the elbow flexors. Exerc. Immunol. Rev. 12, 72–85.
10. Goto S., Radak Z. (2005). Proteins and exercise. In: Mooren, F. C., Volker, K. (Eds.), Molecular and Cellular Exercise Physiology, Human Kinetics. Champaign, USA, 55–71.
11. Nieman D. C., Wentz L. M. (2019). The compelling link between physical activity and the body’s defense system. J Sport Health Sci; 8: 201–17.
12. Radak Z., Ishihara K., Tekus E., Varga C., Posa A., Balogh L., Boldogh I., Koltai E. (2017). Exercise, oxidants, and antioxidants change the shape of the bell-shaped hormesis curve. Redox Biology 12, 285–290.
13. Windsor M. T., Bailey T. G., Perissiou M., Meital L., Golledge J., Russell F. D., Askew C. D. (2018). Cytokine Responses to Acute Exercise in Healthy Older Adults: The Effect of Cardiorespiratory Fitness. Front. Physiol. 9: 203.
14. Fischer C. P. (2006). Interleukin-6 in acute exercise and training: what is the biological relevance? Exercise Immunology Review, 6–33.
15. Leal L. G., Lopes M. A., Batista M. L. (2018). Physical Exercise-Induced Myokines and Muscle-Adipose Tissue Crosstalk: A Review of Current Knowledge and the Implications for Health and Metabolic Diseases. Front. Physiol. 9: 1307.
16. Northoff H., Berg A. (1991). Immunologic mediators as parameters of the reaction to strenuous exercise. Int J Sports Med 12, Suppl, 1: S, 9–15.
17. Larrabee R. C. (1902). Leukocytosis after violent exercise. J Med Res (NS); 7: 76–82.
18. Kishimoto T. (1989). The biology of interleukin-6. Blood 74: 1–10.
19. Kerksick С. М., Wilborn C. D., Roberts M. D., Smith-Ryan A., Kleiner S. M., Jäger R., Collins R., Cooke M., Davis J. N., Galvan E., Greenwood M., Lowery L. M., Wildman R., Antonio J., Kreider R. B. (2018). ISSN exercise & sports nutrition review update: research & recommendations. Journal of the International Society of Sports Nutrition 15: 38.
20. Chycki J., Kostrzeva M., Maszczyk A., Zajac A. (2021). Chronic ingestion of rich-bicarbonate water improves anaerobic performance in hypohydrated elite combat sport athletes. Int J of Env Res and Public Health, 18.
21. Nicholls D. G., Bernson V. S., Heaton G. M. (1978). The identification of the component in the inner membrane of brown adipose tissue mitochondria responsible for regulating energy dissipation Experientia Suppl.,Vol. 32., 89–93.
22. Rupprecht A., Sittner D., Smorodchenko A., Hilse K. E., Goyn J., Moldzio R., Seiler E., Anja U., Brauer A. E. M., Pohl E. E., (2014). Uncoupling Protein 2 and 4 Expression Pattern during Stem Cell Differentiation Provides New Insight into Their Putative Function. PLoS ONE 9(2): e88474.
23. Chaudhuri L., Srivastava R. K., Kos F., Shrikant P. A. (2016). Uncoupling protein 2 regulates metabolic reprogramming and fate of antigen-stimulated CD8+ T cells, Cancer Immunol Immunother. 65 (7): 869–874.
24. Echtay K. S., Winkler E., Bienengraeber M., Klingenberg M. (2000). Site-directed mutagenesis identifies residues in uncoupling protein (UCP1) involved in three different functions. Biochemistry; 39: 3311–3317.
25. Dietrich M. O., Andrews Z. B., Horvath T. L. (2008). Exercise-Induced Synaptogenesis in the Hippocampus Is Dependent on UCP2-Regulated Mitochondrial Adaptation. Journal of Neuroscience 15 October, 28 (42): 10766–10771.
26. Raschke S., Elsen M., Gassenhuber H., Sommerfeld M., Schwahn U., Brockmann B., Jung R., Wisløff U., Tjønna A. E., Raastad T., Hallén J., Norheim F., Drevon C. A., Romacho T., Eckardt K., Eckel J. (2013). Evidence against a Beneficial Effect of Irisin in Humans//PLoS ONE. – September 11 (Vol. 8, № 9), e73680.
27. Raichlen D. A., Klimentidis, Y. C., Bharadwaj, P. K., Alexander G. E. (2019). Differential Associations of engagement in physical activity and estimated cardiorespiratory fitness with brain volume in middle-aged to older adults in Brain Imaging and Behavior.
28. Raichlen D. A., Alexander G. E. (2017). Adaptive capacity: an evolutionary neuroscience model linking exercise, cognition, and brain health. Trends in Neurosciences, Vol. 40, № 7, 408–421.
29. Clark A., Mach N. (2016). Exercise-induced stress behavior, gut microbiota-brain axis and diet: a systematic review for athletes. Journal of the International Society of Sports Nutrition, 13–43.
30. Zeppa S. D., Agostini D., Gervasi M., Annibalini G., Amatori S., Ferrini F., Sisti D., Piccoli G., Barbieri E., Sestili P., Stocchi V. (2020). Mutual Interactions among Exercise, Sport Supplements and Microbiota. Nutrients 2020, 12, 17.
31. Oettlé G. J. (1991). Effect of moderate exercise on bowel habit. Gut, 32, 941–944.
32. Hagio M., Matsumoto M., Yajima T., Hara H., Ishizuka S. (1985). Voluntary wheel running exercise and dietary lactose concomitantly reduce proportion of secondary bile acids in rat feces. J. Appl. Physiol., 109, 663–668.
33. Cerda B., Perez M., Perez-Santiago J. D., Tornero-Aguilera J. F., Gonzalez-Soltero R., Larrosa M. (2016). GutMicrobiota Modification: Another Piece in the Puzzle of the Benefits of Physical Exercise in Health, Front. Physiol., 7, 51.
34. Francaux M. (2009). Toll-like receptor signalling induced by endurance exercise. Appl. Physiol. Nutr. Metab., 34, 454–458.
35. Frosali S., Pagliari D., Gambassi G., Landolfi R., Pandolfi F., Cianci R. (2015). How the Intricate Interactionamong Toll-Like Receptors, Microbiota, and Intestinal Immunity Can Influence Gastrointestinal Pathology. J. Immunol. Res., 2015, 489821.
36. Viloria M., Lara-Padilla E., Campos-Rodríguez R., Jarillo-Luna A., Reyna-Garfias H., López-Sánchez P., Rivera-Aguilar V., Salas-Casas A., Berral de la Rosa F.J., García-LatorreE. (2011). Effect of moderate exercise on IgA levels and lymphocyte count in mouse intestine. Immunol. Investig., 40, 640–656.
37. Turnbaugh P. J., Ley R. E., Mahowald M. A., Magrini V., Mardis E. R., Gordon J. I. (2006). An obesity-associated gut microbiome with increased capacity for energy harvest. Nature, 444, 1027–1031.
38. Mailing L. J., Allen J. M., Buford T. W., Fields C. J., Woods J. A. (2019). Exercise and the Gut Microbiome: A Review of the Evidence, Potential Mechanisms, and Implications for Human Health. Exerc. Sport Sci. Rev. 2019, 47, 75–85.
39. Dokladny K., Zuhl M. N., Moseley P. L. (2016). Molecular Adaptations to Exercise, Heat Acclimation, and Thermotolerance. J. Appl. Physiol. 15; 120 (6): 692–701.
40. Andrews P. W., Thomson J. A. Jr. (2009). The bright side of being blue: depression as an adaptation for analyzing complex problems. Psychol. Rev.; 116 (3): 620–54.
41. Stix G. (2021). Evolution Could Explain Why Psychotherapy May Work for Depression. Scientific American, February, 14.
42. Clark A., Mach N. (2017) The Crosstalk between the Gut Microbiota and Mitochondria during Exercise. Front. Physiol. 8: 319.
43. Lutz H. L., Jackson E. W., Webala P. W., Babyesiza W. S., Kerbis Peterhans J. C., Demos T. C., Patterson B. D., Gilbert J. A., (2019). Ecology and host identity outweigh evolutionary history in shaping the bat microbiome. mSystems, 4: e 00511–19.
Глава V. Цель – все и ничто
Только хорошо поняв второе начало термодинамики, можно ответить на вопрос, почему вообще возможна жизнь.
Карло Черчиньяни (1998), итальянский математик, физик-теоретик, поэт и эссеистЭнтропия к месту и не очень
Разумеется, в эволюционном движении живого, как разновидности движения материи и энергии, нет никакого предопределенного смысла, а тем более замысла – ни скрытого, ни тем более явного, несмотря на то что они «ясно видны» многим «посвященным». Если где-то в данной книге и используются выражения вроде «эволюция сделала вот это, чтобы решить вот то», на самом деле это всего лишь «обытовленное» объяснение динамики генетических ансамблей, к которым «принципы статистической физики применимы столь же, сколь и к ансамблям молекул» (Кунин Е. В., 2012).
Все формы взаимодействия в биологических и протобиологических средах (сложных органических молекул) можно в своих крайних пределах свести к двум формам: отношениям протагонистов или дружественности (сотрудничество, синтрофия, симбиоз, кооперация и т. д.) и отношениям антагонистов или враждебности (прямое соперничество, конкуренция, с одной стороны, и хищничество, паразитизм, зачастую вырастающий из сотрудничества, с другой). Граница между ними тонкая, если не зыбкая; нейтральных же взаимодействий, пожалуй, нет совсем. Соответственно, ответ на вопрос, на каких этапах развития материи и в какой последовательности эти явления возникают, и каким образом они вытекают из оснований статистической физики, может стать и предвестником ответа на вопрос о сущности болезней, воспринимаемых часто как потери совершенства изначально почти безупречных организмов или как прямые следствия их некоего изначального несовершенства. Представляется, однако, что именно несовершенства, как отклонения от совершенного низкоэнтропийного «идеала», реальные или кажущиеся, и делают живой (и не только живой) мир таким, каков он есть: развивающимся от простого к все более сложному, постоянно генерирующим что-то новое, неизбежно забывающим и неизбывно повторяющим (почти) все старое, в котором все в отдельности неизбежно скоро заканчивается и когда-то закончится и все целиком.
В третьей главе на отдельных примерах были кратко рассмотрены квантовые и энтропийные основания базовых энергетических предпосылок жизни и сопутствующая им диссипативность, то есть «набор свойств открытой, неравновесной, рассеивающей энергию системы, движимой градиентом энергии, что увеличивает энтропию окружающей среды». Несомненно, что физические, в первую очередь термодинамические основания, второе начало термодинамики являются наиболее всеобъемлющим оправданием существования всего живого с его общим движением к нарастанию энтропии, но с неизбежным возникновением локальных и преходящих ее снижений (флуктуаций), то есть упорядочиваний хаоса. Рассмотрение энтропии как чистой «меры беспорядка» может запутать: очевидно, что энтропия «покоящейся» системы – ровного песка в пустыне, сбалансированной группы клеток, стагнирующего человеческого общества – гораздо выше, соответственно, энтропии осыпающегося бархана, смеси клеток хозяев и клеток новых паразитов, общества в состоянии революции и гражданской войны, ввиду того, что совокупность или возможность «выбора» доступных состояний для каждой единицы или вариантов совокупностей микросостояний в «спокойной» системе гораздо больше возможностей выбора в состоянии «революции».
Так, лидер партии «Яблоко» Григорий Явлинский в своей статье «Политическая энтропия. Цифровые технологии и глобализация беспорядка» (2020) предполагает, что на смену эпохи глобализации, предположительно, более упорядоченной, приходит «век беспорядка», где «существует множество труднопредсказуемых событий и развилок, и куда, как говорится, кривая выведет, не знает никто». И этот переход с ростом беспорядка предполагается рассматривать как рост «политической энтропии». Однако более правильным будет скорее обратное – падение энтропии: несмотря на возникновение потоков событий с непредсказуемой динамикой, для отдельных людей – совокупности элементов системы – количество вариантов выбора («свободы») на самом деле уменьшается, о чем совершенно справедливо указывает и сам Г. Явлинский в заключение своей статьи, связывая это в первую очередь с добровольным отказом людей от свободы выбора, нежели направленной деятельностью авторитарных лидеров. Отдельный большой вопрос, разумеется, насколько этот выбор на самом деле доброволен, но несомненно, что само уменьшение свободы выбора для подавляющего большинства элементов системы – это несомненно снижение ее энтропии. Тем более актуальное в свете взрывного развития информационных технологий и роста все более детализированной информации о системе. Информации, к сожалению, неравномерно распределяемой, но в итоге дающей уменьшение неполноты информации о системе (см. ниже), что также показывает скорее падение «политической энтропии», нежели ее возрастание. И тем более, что «добровольный отказ от свободы» и рост информационной насыщенности общественного пространства могут оказаться двумя сторонами одной медали.
Термодинамика и информация
Со времен установления принципа Людвига Больцмана (ок. 1875 г.) стало возможным рассматривать классическую термодинамическую и информационную энтропию как практически и теоретически очень близкие, если не равнозначные. В формулировке Макса Планка (ок. 1900 г.) этот принцип как связь средней энтропии системы с вероятностью данного состояния определяется логарифмической зависимостью:
S = kln(Ω),
где S – общая энтропия состояния системы, константа k = = 1,38*10-23 Дж/К, названная Планком постоянной Больцмана, а Ω – статистический вес состояния, являющийся числом возможных равновероятных микросостояний (способов), с помощью которых можно составить данное макроскопическое состояние. Более ранняя формулировка Джозайя Гиббса (1878 г.) для микросостояний, имеющих разную вероятность, устанавливает зависимость в виде суммы произведения вероятности состояний на натуральные логарифмы этих же вероятностей, взятой с противоположным знаком:
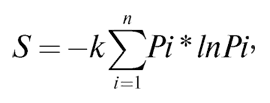
где Pi – вероятность пребывания системы в состоянии i. Минус в начале формулы необходим для приведения обеих частей формулы к общему знаку из-за постоянной отрицательности логарифмов вероятности (так как вероятности всегда меньше 1) и неизменной положительности энтропии. Сам Больцман, комментируя свое физико-статистическое определение энтропии, отмечал, что энтропия характеризует недостающую информацию о системе. В этой интерпретации энтропия может считаться мерой неполноты информации о системе.
В ходе разработки углубленной теории информации информационной энтропией стала считаться мера неопределенности, независимо выведенная как сумма произведений вероятностей событий на логарифмы этих же вероятностей (формула Хартли-Шеннона, основателей теории информации), взятая с противоположным знаком:
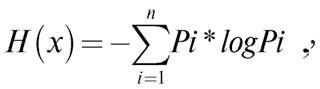
где H(x) – средняя энтропия сообщения, иногда обозначаемая также как мера неопределенности какого-либо события, или даже как количество информации, Pi – вероятность отдельного события i. Основание логарифма в принципе может быть любым больше 1: в двоичных информационных системах – 2, математические статистики, использующие формулу в различных научных дисциплинах, в том числе биологических и клинических, часто используют натуральный логарифм.
Отрицательная энтропия, которую описывал Эрвин Шредингер, это, разумеется, не энтропия с отрицательным знаком – это, очень условно говоря, мера удаленности от максимальной энтропии в рассматриваемой системе, хотя само понятие негативной энтропии (негэнтропии) на самом деле гораздо более сложное, глубокое и весьма запутанное, даже в сравнении с часто неоднозначно понимаемой «обычной», «позитивной» энтропией в своих самых разнообразных интерпретациях. Некоторые исследователи, вслед за Норбертом Винером, увидевшим за очевидным сходством формул Больцмана-Гиббса и Хартли-Шеннона более глубокую связь термодинамической и информационной энтропий, ассоциируют негэнтропию с информацией, что представляется красивым, но до сих пор далеко не всеми признаваемым суждением.
Одной из точек смыкания термодинамической и информационной энтропии являлся, например, известный парадокс «демона Максвелла», гипотетического устройства, стоящего на границе двух объемов и способного различать быстрые и медленные молекулы и, соответственно, пропускать или не пропускать молекулы в одну сторону, что спонтанно ведет к увеличению упорядоченности системы и падению энтропии, что формально противоречит второму началу термодинамики. Решение парадокса было предложено французским и американским физиком Леоном Бриллюэном, математически показавшем, что даже демон ничего не хочет делать бесплатно: чтобы увидеть скорость молекулы, ему нужно заплатить энергией, которая не может быть менее энергии одного теплового кванта (0,025 эВ при комнатной температуре), и эта энергия соответствует минимальной цене одного бита информации. То есть демон совершает работу по обработке информации, и эта работа охлаждает одно тело и нагревает другое, и так же, как за работу любого домашнего холодильника, за эту работу надо платить.

