


Полная версия
Амшены, аргыны, атурая, ашкенази, канглы, их генетические родственники и соседи с древнейших времен до наших дней
Данный вывод подтверждается тем, что неандертальская примесь (2,3%) есть у усть-ишимского человека (село Усть-Ишим Усть-Ишимского района Омской области), жившего около 45 тыс. лет назад [171], у которого определена базальная линия Y-хромосомной гаплогруппы K2a*-M2308 [228].
Неандертальская примесь обнаружена также у человека из пещеры Оасе (Пе́штера-ку-Оа́се (Peştera cu Oase), Румыния, более 40 тыс. лет назад), у которого определена, как и у усть-ишимского человека Y-хромосомная гаплогруппа К2а* [228] и митохондриальная гаплогруппа N [170]. Генетики полагают, что потомков Оасе 1 среди современных европейцев нет [228].

Оасе 1 – человек из пещеры Оасе (Пе́штера-ку-Оа́се (Peştera cu Oase), Румыния, более 40 тыс. лет назад) [227]. Первоначально, исходя из размера челюсти, исследователи предполагали, что Оасе 1 – неандерталец. Анализ ДНК его челюсти, найденной в 2002 г., показал, что у него до 11% неандертальских генов (у современных европейцев таких генов до 4%), что указывает на скрещивание с неандертальцами в предыдущих 4—6 поколениях [228].
У Prüfer K. et al. (2021) приведен возраст женщины из пещеры Златый кунь (Zlatý kůň) (Чехия) – ~43 тыс. лет назад. По мнению исследователей, неандертальские гены предок этой женщины получил за 70—80 поколений до её рождения [230]. Зная возраст женщины, проведем простейшие расчёты, учитывая при этом, что в древности женщины начинали рожать рано, в 12—13 лет. Мы возьмём для расчёта цифру в 15 лет. В этом случае получим: 80х15=1200 лет, 70х15=1050 лет. В итоге получим, что предок женщины, получивший гены от неандертальца, жил примерно 44 тыс. лет назад. В целом, не изменится картина, если в качестве возраста деторождения принять 20 лет: 80х20=1600 лет, 70х20=1400 лет. Время жизни предка женщины – получателя неандертальских генов составит около 45 тыс. лет.
Примечательно, что в этот период на Алтае появились карабомовская (43200±1500 лет назад (49797—44537 кал. лет назад) и 43300±1600 лет назад (49915—44614 кал. лет назад)) [107, с. 293]) и усть-каракольская (около 45 тыс. лет назад) [268]) верхнепалеолитические индустрии. Примерно такой же возраст имеют начальноверхнепалеолитические европейские индустрии богунисьен и бачокирьен, ближневосточный эмиран.
Отметим также, что гаплогруппа, к базальной линии которой относится усть-ишимец, является материнской для K-M2335 (TMRCA 41500 лет назад), от которой произошла гаплогруппа NO [267], предковая для N, преобладающей у уральских народов и О, доминирующей у китайцев и аустронезийских народов. У всех этих народов есть неандертальская примесь.
Поскольку M. Kuhlwilm et al. (2016) утверждают, что алтайские неандертальцы имеют гены современного человека и скрещивание произошло в районе Ближнего (Среднего) Востока 125—120 тыс. лет назад, обратим внимание на Алтай (заметим, что выше мы установили, что на Среднем Востоке (в Передней Азии) гибридизации неандертальцев и современного человека не было).
На Алтае стоянки неандертальцев найдены в трех пещерах: имени Окладникова, Страшной и Чагырской. Следы неандертальцев есть также в Денисовой пещере, где в эту же эпоху обитали денисовцы.
Палеогенетики из Института эволюционной антропологии Общества Макса Планка в Лейпциге (MPI-EVA) выделили ядерную ДНК неандертальцев не из костей, а из пещерного седимента (грунтовых отложений) в Денисовой и Чагырской пещерах на Алтае, а также в Галерии-де-лас-Эстатуас (Galería de las Estatuas (Галерея статуй)) в северной Испании. В этих пещерах, из слоёв возрастом 200—50 тыс. лет, были отобраны 150 образцов пещерного грунта. Для подтверждения верности определения использовали данные ядерной ДНК из костей и зубов и митохондриальной ДНК неандертальцев из тех же отложений (секвенированный геном женщины из Чагырской пещеры, полученный из фаланги пальца, и несколько образцов генома из Денисовой пещеры). В отложениях пещер Галерии-де-лас-Эстатуас и Чагырской обнаружена ДНК только неандертальцев, в Денисовой пещере – денисовцев и неандертальцев, без их смешения.
Геном неандертальцев из Чагырской пещеры существенно отличается от неандертальского генома из Денисовой пещеры. В испанской Галерии-де-лас-Эстатуас присутствуют следы неандертальцев, как из Чагырской, так и Денисовской пещер. По мнению исследователей, это связано с тем, что первая группа неандертальцев мигрировала из Европы в Азию 135 тыс. лет назад, а вторая – 105 тыс. лет назад [259].
Kolobova et al. (2020) установили, что ДНК неандертальцев, обитавших в Чагырской пещере 59—49 тыс. лет назад, свидетельствует об их более тесной связи с восточноевропейскими неандертальцами (Северная Хорватия и Северный Кавказ), чем с неандертальцами из пещеры Денисова, жившими 110 тыс. лет назад. Каменная индустрия из Чагырской пещеры тождественна инструментам микокской культуры эпохи среднего палеолита, распространенной в Восточной и Центральной Европе. Орудия из пещеры Окладникова (Сибирячихинской), расположенной восточнее Чагырской пещеры, имеющие радиоуглеродный возраст от 37,8 до 24,2 тыс. лет, также относятся к микокской культуре. В пещере Денисова, расположенной недалеко от Чагырской, артефактов микокской культуры нет.
Исследователи делают вывод, что неандертальцы приходили на Алтай дважды. Неандертальцы из Чагырской пещеры связаны со второй волной, которая пришла из Европы около 60 тыс. лет назад и принесла технокомплекс микок [195].
В другой работе этой же группы исследователей (Mafessoni F. et al. (2020)) утверждается, что неандертальцы из Чагырской пещеры пришли на Алтай из Европы около 80 тыс. лет назад и принесли специфическую микокскую традицию изготовления каменных орудий. Наиболее близкородственные чагырским неандертальцы обитали в Мезмайской пещере на Кавказе, в которой также нашли микокские каменные орудия. Установлено, что чагырская популяция имела менее 60 представителей и была изолированной [207].
Чагырские неандертальцы (микок) застали на Алтае денисовцев, которые ещё раньше контактировали с более древней волной неандертальцев, которые исчезли за 30 тыс. лет до прихода носителей микокской индустрии на Алтай.
Анализ ДНК девочки, считавшейся денисовкой, показал, что её отец – денисовец12, а мать – неандерталка. Секвенирование генома чагырского неандертальца показало, что материнская линия у девочки представлена «чагырской» волной миграции, а в отцовской линии есть следы древних неандертальцев первой волны миграции.
Исследователи также утверждают, что все современные люди, кроме коренных африканцев, имеют в геноме около 2,4% генов, полученных в результате гибридизации с неандертальцами второй волны, к которой принадлежат хорватские (пещера Виндия) и чагырские неандертальцы. При этом, хорватские неандертальцы внесли больший вклад [207].
Исходя из вышеприведенной информации, можно сделать вывод, что неандертальские гены современные люди получили от неандертальцев с Балкан, создателей микокской индустрии. Однако, поскольку потомки древних людей (или их ближайших родственников) у которых обнаружена неандертальская ДНК, живут в настоящее время на востоке, в Азии, можно предположить, что гибридизация также происходила на территории промежуточной между Балканами и Алтаем. Миграцией с Балкан на восток древних европейцев, также можно объяснить больший процент неандертальской примеси у современных азиатских народов.
1.3. Создатели индустрии раннего и среднего левантийского мустье (Табун D, С) – предки внеафриканского человечества
Левантийское мустье по вариабельности, исходя из стратиграфической последовательности пещеры Табун, подразделяют на три фации – Табун D (раннее левантийское мустье), C (среднее левантийское мустье), B (позднее левантийское мустье.
Табун D
Hershkovitz et al (2018) отмечают, что для индустрии Табун D характерна развитая однополярная леваллуазская13 технология, основанная на производстве удлиненных заготовок (пластин и леваллуазских острий). На ряде стоянок также использовались другие стратегии редукции нуклеусов. Например, на стоянках Рош-Эйн-Мор (Израиль) (около 200 тыс. лет назад), в пещере Мислия на склоне горы Кармель, в 12 километрах к югу от Хайфы (250—150 тыс. лет назад) применялась леваллуа-пластинчатая традиция. Каменные орудия Мислии, выполненные в технике Леваллуа, имеют сходство с орудиями такого типа с африканских стоянок Homo sapiens [186].
Отметим, что существует огромный разброс дат памятников Табун D. Так, К. К. Павленок (2015) пишет, что большинство комплексов мустье типа Табун D (стоянки Эль Коум, Умм-ель-Тлель, Рош Эйн Мор, Эйн Дифла, пещера Хайоним, Мислия и др.) относится к хронологическому интервалу 250—130 тыс. лет назад [95, с.188]. У А. П. Деревянко, М. В. Шунькова, C. В. Маркина (2014) приведена информация – ЭПР-даты индустрии типа Табун D находятся в пределах 230—120 тыс. лет назад, термолюминесцентные14 – 280—230 тыс. лет назад [30, с. 44].
Hershkovitz et al. (2018) останки человека современного анатомического типа из пещеры Мислия (Misliya Cave) датируют возрастом 174±20 тыс. лет назад (календарных) (при стандартном отклонении 2σ), полученным при совместном применении методов урановых рядов (Uranium-series, U-series) и электронно-парамагнитного резонанса (ЭПР) [186]. Однако, Sharp&Paces (2018) утверждают, что у Hershkovitz et al. (2018) нет достоверных U-series дат старше ~ 70 тыс. лет назад и останки имеют минимальный возраст не старше ~ 60—70 тыс. лет [244].
Л. Б. Вишняцкий (2008) приводит следующие даты для памятников Табун D: пещера Мислия – минимальная ОСЛ дата 130±33 тыс. лет назад, стоянка Айн Дифла – ТЛ дата 105±15 тыс. лет назад и ЭПР даты от 102±13 до 162±18 тыс. лет назад, индустрия нижних слоев стоянки Бокер Тахтит – РУ даты (радиоуглеродные15 даты) не более 50 тыс. лет [16, с. 60—61] и отмечает, что материалы «позднего левантийского мустье типа Табун С и В выглядят явно архаичней, чем гораздо более древние материалы мустье типа Табун D» [16, с. 61].
Дате раннего исхода человека современного анатомического типа из Африки не противоречат данные генетики. Так, согласно расчетам YFull, древнейшие в мире гаплогруппы A00 и A0-T, происходящие от «Y-хромосомного Адама», сформировались 235900 (243700—228300) лет назад. При этом, носители базальной линии гаплогруппы A00 проживают в настоящее время в Камеруне, время жизни их последнего общего предка (TMRCA) 37600 (42200—33100) лет назад [267]. Можно полагать, что носители клады A00 никогда Африку не покидали.
Сестринская к кладе A00, макрогаплогруппа A0-T, предковая для всего остального человечества мужского пола, сформировалась 235900 (243700—228300) лет назад, время жизни последнего общего предка (TMRCA) 161300 (169900—152900) лет назад [267]. Расчетный возраст макрогаплогруппы A0-T, почти совпадает с датировками раннего левантийского мустье (Табун D) по Hershkovitz et al. (2018) [186]. Поэтому исходя из возраста гаплогруппы и датировок Табун D можно предполагать, что индустрия Табун D оставлена Homo sapiens и, возможно, носителями гаплогруппы A0-T и её нисходящих линий, вплоть до ВТ (сформировалась 130700 (136400—125100) лет назад), TMRCA 88000 (92500—83600) лет назад) и СТ (сформировалась 88000 (92500—83600) лет назад, TMRCA 68500 (71800—65200) лет назад).
Y-хромосомная гаплогруппа ВТ (если определение выполнено верно), обнаружена у мужчины со стоянки Дольни-Вестонице (Dolní Věstonice; граветт Чехии, около 31 тыс. лет назад) [173]. В Дольни Вестонице определены также Y-хромосомные гаплогруппы СТ (notIJK), F [173] и C1a2 [210] (первоначально была определена как IJK [173]). Предварительно, как ВТ была определена гаплогруппа обитателя стоянки Буран-Кая (BuranKaya3A) в Крыму, жившего 36 тыс. лет назад, но дополнительные производные аллели указывали на то, что это может быть гаплогруппа СТ или С (не C1a2 или C1b). Обитатель Буран-Каи был носителем очень редкой митохондриальной гаплогруппы N1b, имеющей наибольшую концентрацию на Ближнем Востоке и распространенной на территории от Западной Евразии до Африки [150]. Данный факт указывает, что его предки родом с Ближнего Востока. Согласно Ancient DNA: https://ancientdna.info. Map based on public dataset on www.haplogroup.info. (www.indo-european.eu) у BuranKaya3A Y-хромосомная гаплогруппа C-CTS11043 (C1a) и митохондриальная гаплогруппа N1b и представляет он граветтскую культуру [143].
Гаплогруппа ВТ (ВТ-SRY10831.1) определена у двух хуэй (дунгане) [265]. Вполне может оказаться, что это одна из нисходящих линий ВТ (СТ и ниже).
По мнению A.E. Marks (1983), комплексы раннего левантийского мустье существовали вплоть до перехода к верхнему палеолиту – из мустье типа Табун D, продолжавшего развиваться в южных районах Леванта (пустыня Негев, Израиль) до 60—50 тыс. лет назад, происходит эмиран [208]. Л. Б. Вишняцкий полагает, что ввиду значительного хронологического разрыва с ранним левантийским мустье, индустрию стоянки Бокер Тахтит «следует рассматривать, по-видимому, не как продолжение последнего, а как результат вторичного и независимого появления в технологии и типологии переходных комплексов ряда тех же черт, что некогда были свойственны мустье типа Табун D» [16, с.60—61].
С ранним левантийским мустье типа Табун D ассоциируется появление платинчатой микроиндустрии [215] и предполагается связь населения Табун D с распространением леваллуа-пластинчатых индустрий в Евразии [101]. В частности, исследователи отмечают, что пластинчатая микроиндустрия характерна для слоя 23 палеолитической стоянки Кульбулак в отрогах Чаткальского хребта (Ташкентская область, Узбекистан), который предварительно датируется около 180 тыс. лет назад [101, с.68].
А. И. Кривошапкин (2012) считает, что из ближневосточного мустье типа Табун D происходит технология получения пластин с плоскостных и объемных нуклеусов, характерная для Оби-Рахмата (долина реки Пальтау, Бостанлыкский район Ташкентской области) в Узбекистане [70, с.27—28].
По мнению Е. П. Рыбина (2020), появление сходных типичных пластинчатых индустрий начального верхнего палеолита в Леванте и на Алтае является синхронным и, возможно, связано с «развитием на общей основе предковых леваллуа-пластинчатых индустрий Леванта типа раннелевантийского мустье представленных в комплексах Мислия и Рош-эйн-Мор» [107, с.498].
В. Ю. Коен и В. М. Степанчук (2001) прослеживают признаки технокомплекса Табун D, в кременецкой индустрии нижних слоев стоянки Кулычивка в Западной Волыни, датируемых около 31 тыс. лет назад [56, с.90—91]. При этом они отмечают, что нижний слой Кулычивки гетерогенен и представлен орудиями, типичными для технологически сходных индустрий (Богунисьен, Темната слой VI, Эмиран, а также с определенными оговорками леваллуа-мустье типа Табун D), а также ориньякским типологическим компонентом [56, с.91].
К раннему левантийскому мустье типа Табун D относятся, расположенные в горах Южной Осетии пещеры Кударо I, III и Цона (Южная Осетия); пещера Джручула в Имеретии (Западная Грузия); пещера Ховк I и стоянка открытого типа Баграташен I в Северной Армении [29, с.131]. По мнению Э. Ю. Демиденко (2020), возможная инфильтрация людей современного типа (раннее левантийское мустье типа Табун D) на Кавказ, «скорее всего, была эпизодическим и несистемным явлением, так как не привела к передвижению их по всему Северному Кавказу, не говоря уже про иные регионы Восточной Европы, где стоянок данных индустрий среднего палеолита нет» [29, с.133].
В. Б. Дороничев и Л. В. Голованова (2018) также отмечают близость к Табун D мустьерской индустрии кударо-джручульского типа на Южном Кавказе (пещеры Кударо 1, Кударо 3, Цона и Джручула) и исходя из того, что в пещере Джручула найден якобы зуб неандертальца16, считают создателей кударо-джручульской индустрии неандертальцами. Кроме того, они говорят о сходстве с индустриями Южного Кавказа индустрий Северного Кавказа – в бассейне Терека (грот Сарадж-Чуко в Кабардино-Балкарии и пещера Мыштулагты Лагат (пещера Ласок) в Северной Осетии) [41, с.74].
Табун С
Табун С (Схул слой В, Кавзех слой XVII—XXIV, Доудара слой IIIB, Хайоним слой Е, Нар Ибрагим), фиксируется в Леванте в пределах 130—120 тыс. лет назад и существует вплоть до 75 тыс. лет назад [95, с. 188].
Для индустрии Табун С характерно преобладание широких овальных леваллуазских отщепов. С этим типом индустрии ассоциируются находки, как полагают некоторые исследователи, людей современного типа в Кафзех (110—90 тыс. лет назад и Схул17 (130—100 тыс. лет назад) [181; 183].
Нужно отметить, что с периодом существования индустрии Табун С совпадает время формирования Y-хромосомной гаплогруппы ВТ – 130700 (136400—125100) лет назад и время жизни последнего общего предка (TMRCA) её носителей 88000 (92500—83600) лет назад [267].
В случае, если верны даты, приводимые Л. Б. Вишняцким (2008) для памятников Табун D: пещера Мислия – минимальная ОСЛ дата 130±33 тыс. лет назад, стоянка Айн Дифла – ТЛ дата 105±15 тыс. лет назад и ЭПР даты от 102±13 до 162±18 тыс. лет назад [16, с. 60—61], а не Hershkovitz I. et al (2018) – левантийского мустье типа Табун D, верхняя терраса пещеры Мислия – около 194—177 тыс. лет назад) [186], то гаплогруппа ВТ – патрилинейная наследственность представителей левантийского мустье D.
С. В. Дробышевский не считает группу Схул-Кафзех сапиенсами. По его мнению, они «не выходят за рамки видового размаха неандертальцев» и обозначает их P. neanderthalensis palestinensis [45].
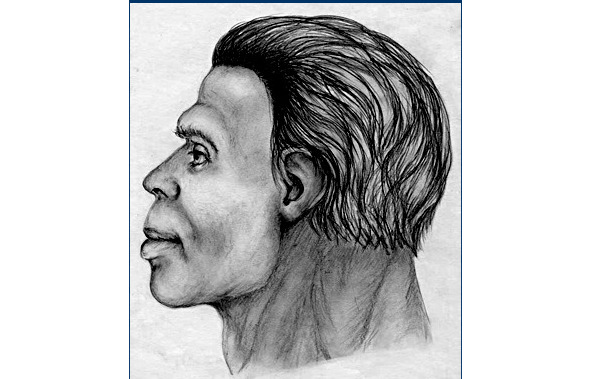

Мужчина из погребения Схул-V. Графическая реконструкция Т. С. Балуевой [125] и изображение в цвете, выполненное по реконструкции художником Philip Edwin [227]
Отметим, что кисть руки Кафзех 9 (Qafzeh 9), имеющего возраст 95 тыс. лет и считающегося ранним современным человеком, отличалась от кисти неандертальцев, тем, что позволяла совершать тонкие и точные движения пальцами. Неандертальцы, в силу особенностей строения кисти, могли хорошо совершать действия с орудиями, имеющими длинные рукояти (копье и др.), но не могли манипулировать с мелкими предметами (вдевать нитки в иголки и т.п.) [144].
К. К. Павленок, Г. Д. Павленок, Р. Н. Курбанов (2020) констатируют, что доминирующей при изготовлении орудий в слое 23 стоянки Кульбулак (Узбекистан) была преференциальная техника леваллуа (preferentiаl Levallois method), распространенная в индустриях типа Табун С (стоянки Табун, Кафзех, Схул слой B, Хайоним слой Е и др.), относящихся к периоду от 190 до 85 тыс. лет назад. Преференциальная техника леваллуа соседствовала «с нуклеусами, расщепление которых велось в рамках параллельного пластинчатого метода, известного в регионе в первую очередь по материалам грота Оби-Рахмат» [101, с.68].
В слое 23 также представлена техника Нар-Ибрагим (Nahr Ibrahim technique), характерная для индустрий типа Табун С, результатом которой являются тронкировано-фасетированные изделия [101, с. 68].
Л. Б. Вишняцкий (2012) выдвинул предположение, что ранние сапиенсы Леванта (группа Схул-Кафзех, датируемая разными методами 81—115 тыс. лет до н.э.), которые, как считается, вымерли после 80 тыс. лет назад, могли мигрировать на юг Аравии, а затем в Австралию [19].
В пользу этой версии имеются археологические данные: комплекс Шибат Дихья I в районе Вади Сурдуд на Аравийском полуострове [162] и антропологические свидетельства. C. P. Groves (1994) указывал на определённое сходство черепа Кафзех 6 с черепами австралийских аборигенов, возможно сохранивших в длительной изоляции древний краниологический комплекс [182]. M. A. Schillaci (2008) проведя многомерный анализ морфометрических характеристик различных человеческих популяций пришёл к выводу, что палеолитическое население Австралии обнаруживает наибольшую генетическую близость к древнейшему населению Леванта. Кроме того, современное и верхнепалеолитическое население Европы имеет общего предка, отличающегося от древних жителей Австралии и древнейшего населения Леванта. Это, по его мнению, свидетельствует о том, что древнейшее население Леванта является непосредственным предком австралийцев-аборигенов или они имеют общего предка [242].
Несостоятельность гипотезы показывает то, что гаплогруппа C1-Z31885, наиболее часто встречающаяся у австралийцев-аборигенов сформировалась 47200 лет назад, время жизни последнего общего предка её представителей (TMRCA) 41100 лет назад. Вместе с тем, прародительская для C-Z31885 гаплогруппа С (сформировалась 65900 (69100—62700) лет назад, TMRCA 48800 (51300—46400) лет назад), является «внучкой» гаплогруппы СТ (сформировалась 88000 (92500—83600) лет назад, TMRCA 68500 (71800—65200) лет назад) и «дочерью» CF (сформировалась 68500 (71800—65200) лет назад, TMRCA 65900 (69100—62700) лет назад) [267]. Поэтому, можно предположить, что носители гаплогрупп СТ и CF внешне были похожи на австралийцев-аборигенов.
Дочерняя для СТ гаплогруппа CF и дочерняя для СF гаплогруппа F (сформировалась 65900 лет назад, TMRCA 48800 (51400—46300) лет назад) [267], судя по времени обитания ранних Homo sapiens в Леванте 250—130 тыс. лет назад [95, с. 188], 130—80 тыс. лет назад [16, с.60—61; 245], сформировались не в Леванте.
Табун В
В Леванте, в финале среднего палеолита 75/70—45 тыс. лет назад, во всех экологических зонах – средиземноморской прибрежной и аридной континентальной, господствовал леваллуазский технокомплекс [88, с. 13—14], который в этом регионе также называется мустье типа Табун В (стоянки Амуд (слой B1- 4), Кебара (слой VI—XII), Ксар Акил (слой XXVI), Тор Фараж (слой С), Тор Сабиха (слой С) и др.) [95, с. 188] и ассоциируется с неандертальцами [151; 223]. Эталонными технокомплексами мустье типа Табун В считаются ассамбляжи слоев X и IX пещеры Кебара. В основании слоя XI Кебары обнаружено погребение неандертальца [24, с.79].
Основой расщепления камня в левантийском мустье В являлась леваллуазская конвергентная техника [212; 231].
В. С. Славинский (2021) полагает, что среднепалеолитическая леваллуазская конвергентная однонаправленная техника для получения острий характерна для слоёв 19—21 грота Оби-Рахмат18 в Узбекистане и оставлены эти слои неандертальцами [116, с.34]. Оби-Рахмат, по его мнению, был промежуточным пунктом на пути неандертальцев из Леванта на Алтай и далее на восток. Индустрии, содержащие артефакты леваллуазской конвергентной техники, фиксируются на территории Алтая в частности и, по всей видимости, Южной Сибири в интервале примерно 59—50 тыс. лет назад [116, с. 31—33].
Заметим, что происхождение алтайских неандертальцев, согласно недавно полученным данным, выводится из Европы [195; 207]. Все современные люди, кроме коренных африканцев, имеют в геноме около 2,4% генов, полученных в результате гибридизации с неандертальцами второй волны, к которой принадлежат хорватские (пещера Виндия) и чагырские неандертальцы. При этом, хорватские неандертальцы внесли больший вклад [207].
1.4. Влияние извержения супервулкана Тоба на популяции современного человека и неандертальцев
Вполне возможно, что неандертальцы появились на Ближнем Востоке, вследствие того, что многие районы на северо-западе Европы в это время стали малопригодными для обитания из-за извержения супервулкана Тоба на острове Суматра (Индонезия), произошедшего 77—69 тыс. лет назад [156; 217]. По результатам аргон-аргонового радиоизотопного датирования получена дата 75,0±0,9 тыс. лет [119].
Кроме Ближнего Востока, неандертальцы в этот период появились в Закавказье (пещеры Джручула и Сакажиа), в Передней Азии (пещера Шанидар), Средней Азии (пещеры Тешик-Таш, Оби-Рахмат, Ангилак), на Алтае (пещера Окладникова) и смежных с ним районах Центральной Азии [17].
Объём продуктов извержения супервулкана Тоба составил не менее 2,8 тыс. км³, из которых 800 км³ выпало в виде пепла. Пепел Тоба обнаружен в Южной Азии (в Малайзии мощность его слоя достигает 9 м, в Индии – 6 м,), в отложениях Южно-Китайского и Аравийского морей и Индийского океана и в Восточной Африке (микроскопическbq слой тефры вулкана Тоба обнаружен в донных отложениях озера Ньяса (Малави)). Следы извержения Тоба обнаружены в Гренландии, где в ледовых кернах, полученных в результате бурения ледового щита острова, фиксируется резкий скачок содержания солей серной кислоты, датированный 74.2±1.7 тыс. лет назад. Аналогичный скачок содержания серы зафиксирован во льдах Антарктиды.
С извержением Тоба связывают наступление вулканической зимы, – общего снижения температуры, вызванного экранирующим эффектом от выброшенных в стратосферу частиц пепла и сернистого газа, продлившейся не менее 6 лет, и очередного пика оледенения, продолжавшегося тысячу лет. Извержение супервулкана на Суматре могло привести к понижению температуры на 4—10° C в Европе, Центральной Азии и Северной Америке. На большей части территории Африки похолодание при этом не превысило 4° C.



