


Полная версия
Амшены, аргыны, атурая, ашкенази, канглы, их генетические родственники и соседи с древнейших времен до наших дней

Амшены, аргыны, атурая, ашкенази, канглы, их генетические родственники и соседи с древнейших времен до наших дней
Рахметолла Рахимжанович Байтасов
Благодарности:
Рахметолла Байтасов
© Рахметолла Рахимжанович Байтасов, 2022
ISBN 978-5-0056-1604-3
Создано в интеллектуальной издательской системе Ridero




ВВЕДЕНИЕ
Без знания истории своих семьи, рода, племени, народа невозможно укреплять духовность и ставить цели в жизни. Поэтому людям свойственно искать свои корни, истоки языка и культуры своей семьи и народа.
В настоящее время к семейными преданиям, архивным документам, добавился новый инструмент, позволяющий находить своих далеких предков – ДНК-генеалогия.
Я знал, что мои предки – казахские баи, репрессированные в 1930-е годы и подавляющее большинство родных и двоюродных братьев моего деда – «врага народа», как и сам дед, погибли на фронте в годы Второй мировой войны. Определив Y-хромосомную гаплогруппу узнал, что я потомок золотоордынского эмира Караходжи, жившего в конце XIV – начале XV вв. При этом, у меня очень редкая гаплогруппа – G1a. Поэтому появилась мысль найти истоки и проследить миграции носителей этой патрилинейной наследственности. Однако оказалось, что это очень трудная, почти невыполнимая задача, поскольку очень мало информации по палеоДНК и современным представителям данной гаплогруппы. Вместе с тем, опираясь на весьма скудные генетические данные и археологические материалы, удалось проследить маршруты миграции моих предков и их генетических родственников, а также ближайших соседей, начиная с эпохи палеолита вплоть до наших дней.
Результаты исследования излагаются в данном труде и будут интересны археологам, генетикам, историкам, любителям генеалогии, истории и этнографии.
Глава I ПРЕДЫСТОРИЯ СОВРЕМЕННОГО ЧЕЛОВЕЧЕСТВА
1.1. Homo bodoensis – непосредственный предок человека современного анатомического типа, Homo sapiens
Международная группа исследователей под руководством палеоантрополога Мирьяны Роксандич (Mirjana Roksandic) из Университета Виннипега (University of Winnipeg) описала Homo bodoensis, являющегося непосредственным предком современных людей. Вид назван по месту обнаружения черепа в 1976г., в долине реки Бодо Д'ар в Эфиопии. Череп принадлежал человеку, жившему в эпоху среднего плейстоцена, около полумиллиона лет назад.

Художественная реконструкция Homo bodoensis (Image credit: Ettore Mazza) [157]
Homo bodoensis обитал в Северной Африке и Юго-Восточной Европе.
В среднем плейстоцене1, 774—129 тыс. лет назад2, в Африке появился человек современного анатомического типа (Homo sapiens), а в Европе неандерталец (Homo neanderthalensis).
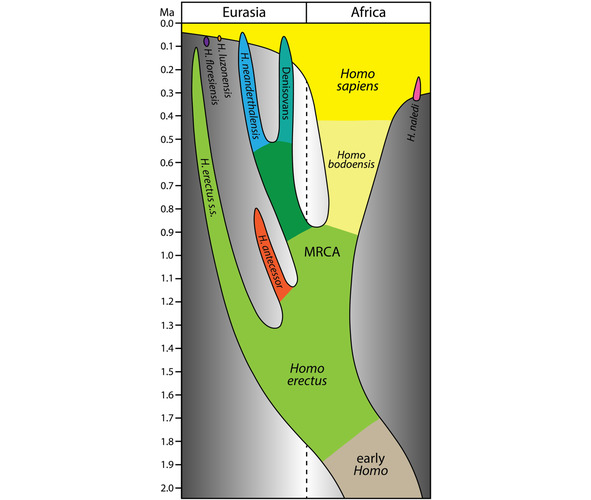
Упрощенная модель эволюции рода Homo в последние 2 млн. лет, в которой Homo bodoensis позиционируется как предок (в основном в Африке) Homo Sapiens [235].
По мнению палеонтологов, к Homo bodoensis следует относить почти все находки Homo, сделанные на африканском континенте и некоторые останки, обнаруженные на юго-востоке Европы.
Останки древних гоминид в Европе, классифицированные, как Homo heldelbergensis и rhodesiensis нужно считать неандертальцами [235].
В 2000-е годы большинство специалистов считало, что Homo sapiens возник в Восточной Африке примерно 200 тыс. лет назад, а современное внеафриканское человечество происходит от группы людей, которая покинула Африку через Баб-эль-Мандебский пролив примерно 65 тыс. лет назад. С территории Южной Аравии люди стали расселяться вдоль побережья Индийского океана на восток [16, c.21].
В 2012 году, генетики, исходя из расчётов скорости мутаций у современных людей, пришли к выводу, что время исхода из Африки следует удревнить до 130—90 тыс. лет назад [239].
Результаты исследования ДНК древних людей из Южной Африки, опубликованные в 2017 году, показали, что Homo sapiens появились на Земле ранее 300 тыс. лет назад [240].
Выход Homo sapiens из Африки, согласно Hershkovitz et al. (2018), мог произойти около 200 тыс. лет назад и ранее – останки человека современного анатомического типа из пещеры Мислия (Misliya Cave) в Леванте имеют возраст, полученный при совместном применении методов урановых рядов (Uranium-series, U-series) и электронно-парамагнитного резонанса (ЭПР)3 174±20 тыс. лет назад (календарных) (при стандартном отклонении 2σ) [186]. Однако, Sharp&Paces (2018) утверждают, что у Hershkovitz et al. (2018) нет достоверных U-series дат старше ~70 тыс. лет назад и останки имеют минимальный возраст не старше ~60—70 тыс. лет [244]. Отметим, что у российских археологов для пещеры Мислия приведена OSL дата4 130±33 тыс. лет назад (предположительно минимальная) [5, c.43].
1.2. Гибридизация человека современного анатомического типа (Homo sapiens) и неандертальцев (Homo neanderthalensis)
1.2.1. Неандертальцы
Неандертальцы (Homo neanderthalensis) – аборигены Европы, имеющие общего предка с человеком современного анатомического типа, жившего примерно 690—555 тыс. лет назад [91]. Согласно Я. В. Кузьмину (2020) возраст наиболее поздних неандертальцев Евразии составляет около 38,8—35,4 тыс. лет назад [80, с.132]. Использование методов ускорительной масс-спектрометрии5 показало, что неандертальцы вымерли около 41030—39260 лет назад [187].

Реконструкция внешности неандертальского мальчика на основании черепа Гибралтар-2 (выполнена специалистами Цюрихского университета с использованием метода компьютерной палеоантропологии) [232]
Рисовали ли неандертальцы?
Несколько лет назад группа британских и немецких исследователей датировала уран-ториевым методом (²³⁴U/²³ºTh)6 кальцитовые натёки на настенной живописи в трех испанских пещерах (Ла-Пасьега (Кантабрияна), Мальтравьесо (Экстремадура) и Ардалес (Андалусия)) и обнаружила, что её возраст – 64,8 тыс. лет. Поскольку в это время в Европе жили только неандертальцы, то по мнению учёных, рисунки оставлены ими [188].

Рисунки в пещере Ла-Пасьега (Испания), около 62 тыс. лет назад [216]
Однако археологи со всего мира усомнились в точности вышеприведенных дат, поскольку уран-ториевый метод мог дать ошибочное определение возраста кальцита7. В случае, если кальцитовые натёки не изолированы от окружающей среды, происходит выщелачивание урана и возраст, полученный уран-ториевым методом удревняется. Например, в пещере Ла-Пасьега для датировки рисунков из более чем 20 полученных дат, была выбрана самая древняя, сильно выбивающаяся из общего ряда и судя по всему, ошибочная из-за выщелачивания урана. Поэтому, по мнению исследователей, нет доказательств способности неандертальцев к рисованию [263].
Не подтверждают умение неандертальцев рисовать, исследования антропологов, которые обнаружили значительные расхождения в анатомии больших пальцев и кистей неандертальцев и современных людей.
Большой палец неандертальцев хорошо вытягивался и отклонялся в сторону, но плохо сгибался внутрь и двигался в стороны. Такая особенность кисти позволяла им крепко держать в руках орудия труда с длинными рукоятками (копьё, молот и т.д.), но затрудняла манипуляции с мелкими предметами, зажимая их между пальцами. Поэтому неандертальцы вряд ли могли вдевать нитки в иголки, хорошо шить [144], можно полагать и рисовать.
При этом, Кафзех 9 (Qafzeh 9), имеющий возраст 95 тыс. лет и считающийся ранним современным человеком, был способен, в отличие от неандертальцев, к более тонким и точным движениям пальцев [144].
Причины вымирания неандертальцев
Наиболее популярны три версии причин, вызвавших вымирание неандертальцев.
1) Сотрудники Института фундаментальных наук IBS (Южная Корея) с помощью суперкомпьютера построили модель, которая называет наиболее вероятной причиной вымирания неандертальцев – проигрыш в конкурентной борьбе за ресурсы с Homo sapiens. Возможными преимуществами современного человека перед неандертальцами называются использование более совершенных методов охоты, более сильный иммунитет, позволяющий противостоять новым заболеваниям и более высокий уровень рождаемости [254].
2) Одна из популярных гипотез – неандертальцы вымерли из-за того, что не смогли приспособиться к резкому изменению климата. Российские исследователи установили, что на Кавказе существует хронологический разрыв между концом среднего палеолита (ассоциируется с неандертальцами) и ранним верхним палеолитом (ассоциируется с Homo sapiens), который совпадает с событием Хейнрих 4, после которого в Кавказском регионе исчезают неандертальцы и появляются высокоразвитые каменные индустрии с преобладанием микропластинчатого компонента, сходные с ахмариеном Леванта [23, с. 171; 25; 26].
По мнению российских исследователей, мощные извержения вулканов, произошедшие практически одновременно на Апеннинах8 и на Кавказе9, привели в Северном полушарии к очень сильному похолоданию, по эффекту сопоставимому с ядерной зимой. Пепел покрывал огромные площади, его слои найдены в восточном Средиземноморье и на Кипре – более чем в 1500 км от извергавшегося вулкана. Следы извержений обнаруживаются в Восточной Европе от Пензы и Ростова на востоке до Южной Украины и Молдовы на западе. Возможно, «площадь поражения» была гораздо больше. Всё это привело к исчезновению неандертальцев [25; 118].
3) Исследование образцов, принадлежавших трём неандертальцам из пещеры Виндия в Хорватии, из Чагырской и Денисовой пещер на Алтае, а также одному денисовцу из Денисовой пещеры, показало, что причиной их вымирания могло стать низкое генетическое разнообразие и возможное распространение гемолитической болезни плода и новорождённого. В свою очередь, низкое разнообразие неандертальцев связано с их географической изоляцией, обусловленной климатическими изменениями в период плейстоцена. Эти факторы в сочетании с конкуренцией с Homo sapiens за пищевые ресурсы в одной и той же экологической нише, могли привести к исчезновению архаичных людей [159].
1.2.2. Время и регионы гибридизации современного человека и неандертальцев
В геномах, как неандертальцев, так и человека современного анатомического типа, обнаружены следы гибридизации друг с другом.
Человеческая ДНК в геномах неандертальцев: источники и время гибридизации
Генетические исследования свидетельствуют, что западноевропейский и кавказский неандертальцы (создатели индустрии восточный микок) имели общего предка около 150 тыс. лет назад [23, с.169].
Изучение ядерной ДНК неандертальцев и денисовцев показало, что они близкие родственники и их предок отделился от предка Homo sapiens 750—600 тыс. лет назад [176]. Установлено также, что неандертальцы и денисовцы скрещивались между собой, а денисовцы, видимо, скрещивались также, с Homo erectus, с которым они контактировали примерно 400 тыс. лет назад [199] и унаследовали от него 1% генома [192].
Сопоставление геномов двух неандертальцев (из Виндии и Денисовой пещеры), денисовца и двух африканцев, показало, что около 3% генома неандертальцев произошло от гибридизации между 200 и 300 тыс. лет назад с древней человеческой группой, ныне вымершей [192].
Сравнение митохондриальной ДНК неандертальца, жившего в пещере Холенштайн-Штадель (Hohlenstein-Stadel) на юго-западе современной Германии около 120 тыс. лет назад, 17 неандертальцев, живших около 430 тыс. лет назад в пещере Сима-де-лос-Уэсос (Sima de los Huesos) в Испании, трёх денисовцев и 54 современных людей, показало, что мтДНК образца из пещеры Холенштайн-Штадель сильно отличается от мтДНК более ранних неандертальцев и показывает сходство с человеческой. Генетики пришли к выводу, что неандертальцы приобрели человеческую митохондриальную ДНК примерно 220 тыс. лет назад, которая к 100 тыс. лет назад почти полностью заменила неандертальскую мтДНК. Исследователи предполагают, что популяция современных людей, вступившая в контакт с неандертальцами, впоследствии вымерла. МтДНК ранних неандертальцев сходна с таковой денисовцев [175; 176; 229].
Учёные также исследовали Y-хромосомы двух денисовских мужчин из Денисовой пещеры (Алтай, Россия), трёх неандертальцев из пещер Эль-Сидрон (Испания), Мезмайской (Кавказ, Россия) и Спи (Бельгия) и сравнили её с Y-хромосомой людей современного типа – африканцев и неафриканцев, представленных геномом человека из Усть-Ишима (Омская область), жившего 45 тыс. лет назад. Исследование показало, что в период от 350 до 150 тыс. лет назад произошла гибридизация неандертальцев с Homo sapiens, в результате чего неандертальцы получили от вымершей ветви человека современного анатомического типа Y-хромосому, которая заместила у них «родные» неандертальские варианты [226].
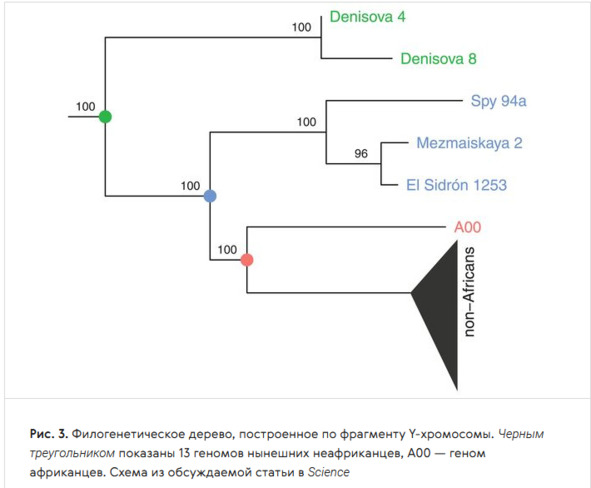
Филогенетическое дерево, построенное по фрагменту Y-хромосомы. Чёрный треугольник – 13 геномов современных неандертальцев; А00 – геном африканцев (Y-хромосомная гаплогруппа А00) [226]
Источники ДНК денисовцев у человека современного анатомического типа
В геноме жителей островов Меланезии (4—6% генома), а также в геномах популяций тихоокеанских островов, китайцев и японцев, есть следы гибридизации с денисовцами [160]. Больше всего денисовской примеси у папуасов. Эта примесь получена ими в результате гибридизации с дальними родственниками денисовцев, живших в Денисовой пещере на Алтае. Возможно, эта популяция денисовцев обитала на юге или юго-востоке Азии. Денисовская примесь есть также у жителей Южной Азии (индусы, бенгальцы и др.). Источник примеси у них тот же, что и у папуасов. Очень незначительная денисовская примесь есть у финнов [249]. Обнаружена денисовская примесь у исландцев, которая, по мнению исследователей, могла попасть к предкам исландцев от неандертальцев, до того, контактировавших с денисовцами либо далекие предки исландцев сталкивались с денисовцами [250].
У китайцев и японцев денисовская примесь получена из двух источников. Один из них, тот же, что и у папуасов, второй – денисовцы близкие к популяции, обитавшей в Денисовой пещере [154].
Время гибридизации современного человека и неандертальца
У европейцев приблизительно 2,5% [258], около 2,4% [207] генома имеет неандертальское происхождение.
M. Kuhlwilm et al. (2016) сравнив геномы неандертальцев, денисовского человека и современных людей, утверждают, что 125—120 тыс. лет назад группа людей современного анатомического типа вышла из Африки в направлении территории Среднего (Ближнего) Востока и Аравийского полуострова, где, по-видимому, обитали неандертальцы. В конечном итоге эта группа людей вымерла, но часть их генома унаследовали неандертальцы, которые мигрировали на Алтай. В геномах денисовца и двух неандертальцев (из Хорватии и Испании) следов ДНК древних африканцев нет, т.е. они не скрещивались с человеком современного анатомического типа.
60—50 тыс. лет назад другая группа людей современного анатомического типа вышла из Африки и встретилась с неандертальцами, мигрировавшими из Европы на Ближний (Средний) Восток и её представители спаривались с неандертальцами. Кроме того, потомки людей из этой группы скрещивались с денисовцами, ДНК которых обнаружена у меланезийцев и китайцев [199].
Chen et al. (2020), исследовав выборку из 2504 человек, происходящих из разных географических регионов, утверждают, что скрещивание современного человека с неандертальцами произошло один раз. Анализ показал, что у всех 504 исследуемых африканцев, неандертальское наследие совпадает с таковым у европейцев, а не у азиатов. Это соответствует сценарию миграции из Европы в Африку какой-то группы европейцев, после того, как их предки отделились от азиатов. Миграция, по-видимому, произошла в последние 20 тыс. лет. В целом, полученные результаты указывают на то, что скрещивание современного человека и неандертальцев происходило однократно, после выхода из Африки [155].
По мнению Browning et al. (2018), гибридизация современного человека с неандертальцами была один раз, либо повторная гибридизация, особенно в Азии, происходила с той же популяцией неандертальцев, с которой человечество столкнулось около 55 тыс. лет назад. Второй вариант более вероятен, так как у народов Азии достоверно больше неандертальской ДНК [154].
Vernot&Akey (2014) полагают, что неандертальцы и люди современного типа скрещивались в период между 75 и 40 тыс. лет назад [258].
С. В. Дробышевский (2019) исходит из данных по усть-ишимскому человеку, у которого неандертальская примесь составляет примерно 2,3%. Момент метисации оценивается в 13—7 тыс. лет до рождения усть-ишимского человека. Учитывая возраст самого усть-ишимца в 45 тыс. лет, смешение должно было происходить 58—52 тыс. лет назад [44]
Регион гибридизации неандертальцев и современного человека и представители Y-хромосомных гаплогрупп Homo sapiens, которые скрещивались с неандертальцами
Л. В. Голованова и В. Б. Дороничев (2010) доказывают, что в Европе неандертальцы и современный человек не скрещивались, поскольку неандертальцы вымерли в этой части света после 40 тыс. лет назад. Они отмечают, что самые поздние радиокарбоновые даты, указывают на существование неандертальцев в Европе незадолго до 38 тыс. лет назад – Фельдхофер в Германии, Эль Сидрон в Испании, Роше-де-Вильнев во Франции, Кюльна в Чехии, слой G3 в Виндии, Хорватия. Более поздние даты являются ошибочными (останки неандертальцев из слоя G1 в Виндии (29080±400 и 28020±360 радиокарбоновых лет) и слоя 3 в Мезмайской (29195±965 радиокарбоновых лет). В Мезмайской пещере возраст слоя 3 с находкой неандертальца определен на основании ЭПР-дат в интервале 73—63 тыс. лет назад. В пещере Виндия, «в слое G1, который формировался в течение длительного времени, вероятно, произошло смешение (палимпсест) стоянок неандертальцев и современных людей10» [24, с. 95].
Вместе с тем, результаты исследования палеоДНК древнейших европейцев, показывают наличие в их геномах ДНК неандертальцев и отсутствие связи с современным населением этой части света. Так, итоги исследования обитателей пещеры Бачо Киро (Болгария), живших 42,5—46 тыс. лет назад, у которых обнаружены следы гибридизации с неандертальцами за шесть-семь поколений до их рождения, свидетельствуют об отсутствии преемственности между создателями индустрии бачокирьена и современными европейцами. Гаплогруппы С1 (C-F3393) и F (F-M89), определенные у них, в настоящее время очень редки и встречаются с низкой частотой лишь в Юго-Восточной Азии и Японии [184].
Расшифровка генома современного человека, череп которого найден в пещере Златый кунь (Zlatý kůň) (Конепрусские пещеры, Чехия), показала, что это женщина, представительница митохондриальной гаплогруппы N. Жила она ~ 43 тыс. лет назад (~43 ka (95% highest posterior density = 31.5—52.6 ka)). Неандертальские гены предок этой женщины получил за 70—80 поколений до её рождения. Женщина из Zlatý kůň генетической преемственности с насельниками Европы, жившими после 40 тыс. лет назад, не имеет. Исследователи считают, что вероятной причиной этого может быть мегаизвержение Флегрейских полей и делают вывод, что большая часть неандертальских генов современным человеком получена в результате смешения с неандертальцами, родственными будущим европейским неандертальцам, а не обнаруженным на Горном Алтае [230].

Женщина из пещеры Златый кунь (Zlatý kůň) (Конепрусские пещеры, Чехия) [227]
S. Condemi et al. (2021), исследовав геномы неандертальцев, денисовца, папуасов, австралийцев-аборигенов и др., обнаружили, как они полагают, свидетельства гибридизации между левантийскими неандертальцами и Homo sapiens около 65 тыс. лет назад. По их мнению, гибридизация произошла во время раннего расселения Homo sapiens через Аравийский полуостров в направлении Австралии и Океании [159].
В исследовании M.A. Almarri et al. (2021) установлено, что в популяциях Ближнего Востока доля генетического вклада неандертальцев, в целом ниже, чем в популяциях Европы и Восточной Азии. По мнению исследователей, это обусловлено двумя причинами: неандертальские фрагменты размывались в ближневосточных геномах из-за смешения с африканской линией и из-за смешения с линией базальных евразийцев11 (те и другие вовсе не имеют неандертальских генов). По этим двум причинам аравийские популяции имеют меньшую долю неандертальского вклада, чем левантийские [140].
Из результатов исследования M.A. Almarri et al. (2021) можно сделать ещё один вывод – гибридизации неандертальцев и современного человека на Ближнем Востоке не было. Косвенно данный вывод подтверждается тем, что в «пещере Кзал-Акир позднемустьерские и древнейшие верхнепалеолитические слои разделены седиментационной или эрозионной лакуной. Позднее мустье таких левантийских памятников, как Кебара или Тор-Фарадж, сильно отличается от древнейших комплексов верхнего палеолита» [82, с. 6].
Если исходить из того, что гибридизация неандертальцев и сапиенсов произошла около 55 тыс. лет назад [154], то с неандертальцами контактировали представители Y-хромосомных гаплогрупп С (сформировалась 65900 лет назад, время жизни последнего общего предка (TMRCA) 48800 лет назад) [267] и F (сформировалась 65900 лет назад, TMRCA 48800 лет назад) [267].
Гибридизация носителей гаплогруппы C с неандертальцами подтверждается тем, что представили её дочерних ветвей С1 и С2, имеют признаки смешения с этим древним человеком. В частности, нандертальская примесь обнаружена у носителей Y-хромосомной гаплогруппы С1 [248], живших в Сунгири (г. Владимир, Россия) 34654—33678 лет назад (калиброванная дата) [114], в Костёнках в Воронежской области России (геном человека Костёнки-14 (возраст 33250±500 некал. лет, 35—36 кал. тыс. лет [112, с.48]) содержит примерно 2,8% неандертальской примеси) [243] и др.
С гаплогруппой F не все так просто, поскольку существуют так называемые базальные евразийцы, у которых нет генов неандертальцев.
Jones E.R. et al. (2015) предполагают, что к базальным евразийцам (Basal Eurasian) относятся кавказские охотники-собиратели (Caucasus hunter-gatherer (CHG)) [194]. Их представители – люди из пещер Сацурблия (возраст 13300 лет; поздний верхний палеолит) и Котиас Клде (возраст 9895—9529 лет; эпоха мезолита). У мужчины из пещеры Сацурблия определены гаплогруппы – Y-хромосомная J1 [267] и митохондриальная К3; у мужчины из пещеры Котиас Клде (Kotias Klde cave) определены гаплогруппы – Y-хромосомная J2а и митохондриальная H13c. В настоящее время эти гапогруппы с высокой частотой встречаются в Грузии [194].
Вопрос о базальных евразийцах будет рассмотрен в отдельном разделе подробнее. Здесь же замечу, что базальными евразийцами, скорее всего, были носители Y-хромосомной гаплогруппы G. Тем не менее, поскольку Jones E.R. et al. (2015) считают обитателей пещер Сацурблия и Котиас Клде базальными евразийцами, можно предполагать, что у древних представителей гаплогруппы J также не было неандертальской примеси.
Определим, носители какой Y-хромосомной гаплогруппы контактировали с неандертальцами.
Гаплогруппа G – «внучка», а J – «прапраправнучка» гаплогруппы F. Материнской для J является гаплогруппа IJ, которая в свою очередь происходит из гаплогруппы IJK.
Поскольку IJ – материнская для гаплогруппы J, носители которой в древности не имели неандертальской примеси, то её представители, можно полагать, не заводили с неандертальцами «семейных» связей. Соответственно, можно предполагать, что с неандертальцами скрещивались представители Y-хромосомной гаплогруппы К (сформировалась 47200 (49700—44600) лет назад, TMRCA 45400 (49600—41400) лет назад), сестринской для IJ. При этом, носители гаплогруппы К должны были проживать в одном регионе с представителями гаплогруппы С (сформировалась 65900 (69100—62700) лет назад, TMRCA 48800 (51300—46400) лет назад) [267].



