Полная версия
Эволюция. Классические идеи в свете новых открытий



Резко уменьшить проходимость ландшафта приспособленности – превратить его в лабиринт узких тропок или горных хребтов – может эпистаз. Так называют взаимное влияние разных мутаций (аллелей) на вредность или полезность друг друга. Замена одной аминокислоты может менять эффективность работы белка не сама по себе, а в зависимости от комбинации других аминокислот, занимающих определенные позиции в том же белке. Также эффект мутации может зависеть от других белков, находящихся в непосредственном взаимодействии с белком-мутантом. Иными словами, для определения смысла биологического текста важен контекст. В этом и состоит суть эпистаза. Мы еще не раз встретимся с этим явлением в последующих главах.
Наличие эпистаза предполагает, что многие аминокислотные замены являются допустимыми только в определенном окружении. Если окружение (контекст) неподходящее, то данная замена будет снижать приспособленность, и отбор ее выметет. Это явление в эволюции белков изучалось ранее на конкретных примерах[22]. Если эпистаз широко распространен, белок может добраться до многих потенциально достижимых пунктов в пространстве последовательностей только долгим обходным путем.
Для проверки гипотезы о действенности эпистаза в эволюции белков Поволоцкая и Кондрашов придумали хитроумный тест, основанный на сопоставлении темпов накопления «сближающих» и «разводящих» мутаций с дистанциями между белками. Основная идея состояла в том, что гипотезы о наличии и отсутствии эпистаза дают противоположные предсказания о частоте закрепления сближающих и разводящих мутаций. Если влияние эпистаза невелико, то у двух близких белков темп накопления разводящих мутаций должен быть высоким, а по мере расхождения последовательностей он должен снижаться (если ландшафт приспособленности легкопроходимый, то белки сначала свободно и быстро «блуждают» по ровному плато, соответствующему данной функции, уходя все дальше от исходной точки, но по мере приближения к его краям их расхождение замедляется). Темп накопления сближающих мутаций при этом должен оставаться примерно постоянным. Если же эпистаз оказывает сильное влияние на эволюцию белков, все должно быть наоборот: разводящие мутации должны накапливаться с примерно постоянной скоростью, а темп накопления сближающих мутаций по мере расхождения белков должен снижаться.
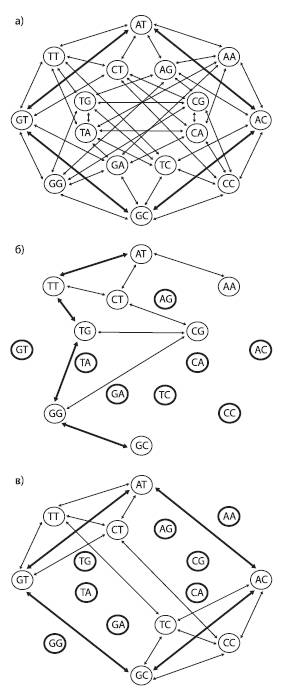
Пространство последовательностей можно изобразить в графа, вершины которого соответствуют разным последовательностям (в данном случае показаны последовательности из двух нуклеотидов), а ребра – единичным эволюционным событиям (нуклеотидным заменам). В первом случае (a) ландшафт приспособленности представляет собой сплошное ровное плато: все 16 последовательностей имеют одинаково высокую приспособленность и все возможные мутации разрешены (не будут отбракованы отбором). Два кратчайших пути, соединяющие последовательности AT и GC, состоят всего из двух мутационных шагов (жирные стрелки). Во втором случае (б) половина последовательностей имеют пониженную приспособленность (отмечены жирными кругами). При этом снижение приспособленности определяется не конкретным нуклеотидом в конкретной позиции, а уникальной комбинацией обоих нуклеотидов (т. е. имеется эпистаз). Это резко снижает проходимость ландшафта: число доступных траекторий уменьшается и кратчайший путь между двумя пунктами (например, от AT к GC) удлиняется. В третьем случае (в) тоже половина последовательностей имеют пониженную приспособленность, но эпистаза нет: нуклеотиды A и G во второй позиции снижают приспособленность независимо от состояния первой позиции. Отсутствие эпистаза способствует тому, что ландшафт остается легкопроходимым, и от AT к GC можно прийти всего за два шага, как и в первом случае. По рисунку из Povolotskaya, Kondrashov, 2010.
Проведенные расчеты подтвердили гипотезу о сильном влиянии эпистаза на эволюцию белков. Оказалось, что темп накопления разводящих мутаций не зависит от дистанции между белками. В каждый момент времени лишь около 2 % аминокислотных позиций могут быть изменены без снижения приспособленности, хотя в долгосрочной перспективе более 90 % позиций могут измениться – но к этим изменениям нужно идти долгими обходными путями по лабиринту узких «горных хребтов» ландшафта приспособленности. У близких, недавно разошедшихся белков темп накопления сближающих мутаций очень высок, потому что у близких белков аминокислотный «контекст» для каждой позиции является сходным. Поэтому та аминокислота, которая недавно стояла в данной позиции, с большой вероятностью может «вернуться» на свое место, и это не снизит приспособленность. Напротив, у сильно различающихся белков данная позиция уже находится в разных контекстах, и поэтому «возвращение» аминокислоты, которая стояла здесь у далекого предка, скорее всего, снизит приспособленность, и мутация будет отсеяна.
В снижении вероятности сближающих мутаций по мере расхождения белков проявляется правило необратимости эволюции. Чем сильнее разошлись белки, тем меньше у них шансов снова стать похожими (мы еще вернемся к этому правилу в главе 5).
Таким образом, ландшафт приспособленности древних белков отличается повышенной «складчатостью». Он похож на сложный лабиринт узких горных хребтов, передвигаться по которому эволюционирующие белки могут лишь очень медленно. Мало того, некоторые узкие мостики норовят обрушиться в бездну после того, как по ним «прополз» эволюционирующий белок, – совсем как в голливудских боевиках, – что придает эволюции элемент необратимости.
Между прочим, полученные результаты – еще один аргумент в пользу единства происхождения всего живого. Подобно тому как разбегающиеся галактики указывают на существовавший в прошлом единый центр, из которого все объекты во Вселенной начали свое движение, так и продолжающееся «разбегание» древних белков указывает на их происхождение от единого предка.
Глава 2. Полезные ошибки
Все здание теории эволюции, а значит и биологии в целом, основано на том, что некоторые случайные наследственные изменения оказываются полезными, т. е. повышают эффективность размножения (по-другому называемую приспособленностью). Термин «полезные мутации» означает только это. В эволюционной биологии бессмысленно говорить о пользе, не имея при этом в виду размножение. Если организм-мутант лучше подлаживается к своей среде, лучше себя чувствует и дольше живет, но при этом проигрывает в размножении, то мутация, какой бы полезной она ни казалась, является вредной. Обычно все эти вещи коррелируют друг с другом, но не всегда.
Для биолога существование полезных мутаций – самоочевидный, банальный факт. Однако многие непрофессионалы склонны усматривать здесь парадокс. Ведь мутации – это ошибки, сбои в системе копирования наследственной информации. Из повседневного опыта мы знаем, что случайные ошибки ничего не улучшают, а только портят. Разве можно улучшить текст, внося в него случайные ошибки?
Это рассуждение верно, если текст совершенен. Действительно, не будем же мы подправлять «Евгения Онегина» или кое-что переписывать в «Женитьбе Фигаро». А если текст черновой и небрежный? Тогда можно и исправить, и улучшить, в том числе путем внесения случайных изменений с отбором удачных вариантов.
Рассмотренная в главе 1 модель с эволюционирующими фразами показывает, как постепенно текст становится лучше. Если нет даровитого цепкого редактора, то особенно важно, чтобы текст тиражировался во множестве копий. В каждую из них вносятся разные ошибки, и для последующего тиражирования раз за разом отбираются лучшие варианты.
В отличие от «Евгения Онегина» биологические тексты редко бывают совершенными[23]. К тому же мир имеет обыкновение меняться. То, что было превосходным вчера, сегодня уже устарело и никуда не годится. Если вчера пределом мечтаний считалось обладание игровой приставкой, то сегодня продвинутый подросток должен иметь последнюю модель айпада (кое-кто из отставших и устаревших уже путают айпад и айпод). Так и в мире природы – меняется биологическое окружение, климат, очертания континентов, и нужно постоянно им соответствовать… Правда, в природе перемены обычно происходят медленнее, чем в современной моде и технологиях, но из этого правила тоже есть исключения.
Итак, мутации – случайные изменения ДНК – бывают полезными. Процесс приспособления неостановим и бесконечен. Но общие рассуждения, как бы громко, уверенно и веско они ни были высказаны, не убедят нашего вдумчивого и скептического читателя. Поэтому постараемся показать на простых примерах, что полезные мутации – это повседневная реальность. В этой главе мы познакомим читателя с несколькими ясными случаями.
Один верный шаг – и на полях вырастает рис
Ключевым моментом в доместикации риса (как и многих других злаков) было появление разновидности с неопадающими семенами. Это позволило древним земледельцам не подбирать зрелые зерна с земли, а срезать их со стеблями и сократить потери при сборе урожая. Предком культурного риса (Oryza sativa) был дикорастущий вид с опадающим семенами O. nivara или близкий к нему O. rufipogon (или, возможно, оба этих вида).
Опадение семян зависит от гена sh4. Для дикого риса O. nivara характерен доминантный аллель[24] этого гена, соответствующий опадающим семенам. Очевидно, что в дикой природе полезны именно опадающие семена. Рецессивный аллель, характерный для домашнего риса, обеспечивает неопадающие семена. В 2006 году ученым из Мичиганского университета (США) удалось выяснить, какую функцию выполняет кодируемый этим геном белок и как он контролирует опадение семян (Li et al., 2006). Обычно в том месте, где должно произойти отделение (например, спелых плодов или отслуживших листьев), формируется «отделительный слой» из клеток особого строения. У дикого риса между зерном и плодоножкой тоже образуется отделительный слой, состоящий из мелких тонкостенных клеток. У культурного риса этот слой формируется только частично, поэтому зерно остается прочно прикрепленным к плодоножке. Так вот, выяснилось, что ген sh4 экспрессируется там и только там, где формируется отделительный слой. Значит, sh4 участвует в организации функционального отделительного слоя. У культурного риса его функциональность нарушилась, и рис приобрел нужные земледельцу свойства.
В последовательности этого гена, расположенного на четвертой хромосоме, удалось определить нужную мутацию. Оказалось, что это одна-единственная нуклеотидная замена: тимин (T) дикого риса заменился на гуанин (G) домашнего, что привело к замене аминокислоты лизина на аспарагин в соответствующем белке. Неужели всего одна аминокислота – и такой чудесный результат? Именно так. И чтобы не было сомнений – а они всегда остаются, пока не выполнишь проверку или не заглянешь в ответы в конце задачника, – ученые ввели в геном культурного риса «дикий» вариант гена sh4. У культурного риса созрели колосья с неудобными, опадающими семенами.
Исследование показало, что доместикация риса была связана с отбором растений, несущих мутантный вариант гена sh4. Мутация немного «подпортила» механизм формирования отделительного слоя, хотя и не вывела его из строя полностью, иначе отделительный слой не формировался бы вовсе, что создало бы большие проблемы при обмолоте. По-видимому, небольшие изменения регуляторных генов сыграли важную роль и в доместикации других растений. Соответствующие данные уже имеются, например, по кукурузе и томатам.
Итак, вот первый конкретный пример полезной мутации. Кстати, почему мы говорим, что она была полезной? Для дикого риса мутация, не позволяющая созревшему зерну своевременно отделиться от колоса, была бы однозначно вредна – она снизила бы число посеянных семян, т. е. снизила эффективность размножения. Но «ландшафт приспособленности» изменился для риса, когда его размножение (сбор урожая и сев) взяли в свои руки земледельцы. Теперь семена, прочно держащиеся в колосе, получили больше шансов добраться до мест обмолота, а значит, быть посеянными и дать потомство. Бессознательный отбор, осуществлявшийся ранними земледельцами, дал преимущество мутантным растениям с неопадающими семенами, т. е. сделал растения с этим признаком более приспособленными по сравнению с «диким типом» – растениями с опадающими семенами. Это, собственно, и означает, что мутация, мешающая семенам опадать и вредная для диких злаков, стала полезной для тех растений, размножение которых люди взяли в свои руки.
Можно также заметить, что новый полезный признак – между прочим, признак важный, составляющий главное отличие культурного злака от дикого, – на молекулярном уровне был связан не с созданием чего-то нового, а всего лишь с повреждением, порчей чего-то старого. Мутация повредила механизм формирования отделительного слоя. Это соответствует общепринятому – и в целом верному – представлению о том, что случайные мутации, как и любые «случайные ошибки», с большей вероятностью могут испортить что-то сложное, чем создать. Однако, как видим, повреждение на молекулярном уровне не обязательно сопровождается упрощением на уровне организма. Оно может приводить к появлению новых полезных признаков и обеспечивать адаптацию к меняющимся условиям. Эволюция, как всякий вероятностный процесс, обычно выбирает самые простые (вероятные) пути из всех возможных. Если можно улучшить приспособленность, всего лишь упростив систему (например, выведя из строя какой-то ген), то именно это, скорее всего, и произойдет[25].
Кто-нибудь может подумать, что так просто все получается лишь с искусственным отбором, если в дело вмешивается человек со своим разумением и целеполаганием. Это не так. Есть превосходные примеры полезных мутаций и среди природных объектов. Скажем, таких, как американские белоногие хомячки.
Десять тысяч лет, три нуклеотида и защитная окраска
Американские белоногие хомячки, называемые также оленьими мышами (Peromyscus maniculatus), в норме имеют темную окраску, однако представители этого вида, обитающие в районе с очень светлой почвой (Песчаные Холмы в штате Небраска), окрашены светлее своих сородичей. Хомячки с Песчаных Холмов стали хрестоматийным примером эволюции адаптивных признаков. Этому способствовали два обстоятельства. Во-первых, адаптивность (полезность) светлой окраски не вызывает сомнений: хищные птицы гораздо лучше видят на светлом фоне темного хомячка, чем светлого. Во-вторых, Песчаные Холмы – молодое геологическое образование: они сформировались после отступления ледника около 10 тыс. лет назад. Это дает основания думать, что мы имеем дело с адаптацией, появившейся – в эволюционном масштабе времени – совсем недавно. Однако для понимания механизмов адаптации важно выяснить ее генетические основы.
Известно, что окраска шерсти млекопитающих зависит от распределения двух пигментов: черно-коричневого эумеланина и рыжего феомеланина. Клетки меланоциты, отвечающие за окраску волоса, могут поочередно синтезировать то один, то другой пигмент по мере роста волоса. В результате волос получается неоднородно окрашенным. Часто кончик и основание волоса темные (эумеланиновые), а посередине имеется светлая (феомеланиновая) полоса. И вот любопытно, что же определяет разницу в окраске у темных и светлых хомячков? Если вглядеться попристальней, то окажется, что никаких специальных ухищрений и особой цветовой палитры не требуется. Всего лишь нужно в каждой шерстинке светлых хомячков расширить феомеланиновую полосу. И больше ничего! Элементарно! И вот американские генетики из Гарвардского и Калифорнийского университетов решили изучить, что же стоит за этим элементарным решением (Linnen et al., 2009).
Известно, что на окраску шерсти может влиять ген Agouti. Сигнальный белок, кодируемый этим геном, подает команду меланоцитам синтезировать феомеланин вместо эумеланина. Влияние мутаций Agouti на окраску шерсти подробно изучено на домовых мышах (Mus musculus). Оказалось, что мутанты с отключенным геном Agouti имеют черную окраску, а повышенная активность гена приводит к очень светлой окраске. Светлый аллель – доминантный по отношению к темному.
Чтобы проверить, действительно ли светлая окраска хомячков с Песчаных Холмов определяется доминантной мутацией, ученые скрещивали разномастных родителей. Все потомство от скрещивания получилось светлым, и эта гипотеза подтвердилась.
Дальнейшие эксперименты показали, что непосредственным результатом мутации является повышенная активность Agouti в первую неделю жизни хомячат, т. е. в тот период, когда у них отрастает шерсть. Максимальная активность Agouti приходится на четвертый день жизни как у темных, так и у светлых хомячков, однако абсолютная величина этой активности (измеряемая по количеству матричных РНК, считанных с гена) у светлых особей с Песчаных Холмов в несколько раз выше.
После была проведена весьма трудоемкая и кропотливая работа – секвенирование нуклеотидной последовательности Agouti у сотни хомячков из темных и светлых лабораторных линий, а также у диких зверьков из зоны смешения темной и светлой популяций. В результате было выявлено около 20 полиморфных сайтов, т. е. таких участков гена, которые не у всех особей одинаковы. Существуют эффективные методы статистического анализа нуклеотидных последовательностей, которые позволяют обнаруживать следы действия положительного отбора на те или иные участки генов. Иными словами, отличать изменения ДНК, которые поддерживались отбором (т. е. полезные, адаптивные), от нейтральных изменений, которые распространялись в популяции за счет дрейфа. Применив эти методы, ученые пришли к выводу, что ключевая мутация, в результате которой мыши приобрели светлую окраску, находится только в одном месте, а изменения в остальных 19 полиморфных сайтах тоже внесли свой вклад, но менее значительный.
Эта мутация находилась лишь в одном из 20 сайтов. Она состояла в выпадении трех нуклеотидов, кодирующих аминокислоту серин. Удивительно, что такой важный признак, как маскирующая окраска, может определяться всего тремя нуклеотидами! Какая поразительная несоразмерность: с одной стороны, три молекулы, их даже в микроскоп не видно, а с другой стороны – невидимость для врага, долгая жизнь, многочисленная семья и потомство… Когда сравниваются такие категории, то кажется, что три молекулы – это ничто, не могут они определить хомячьего счастья. Но эта несоразмерность – мнимая.
Следы естественного отбора
Влияние естественного отбора на генофонд популяции трудно наблюдать в природе из-за медленности процесса. Непосредственно наблюдать отбор, действовавший на популяцию в прошлом, и вовсе невозможно без машины времени. К счастью, это и не обязательно, потому что отбор, нацеленный на тот или иной локус (участок молекулы ДНК), оставляет в геномах легкоразличимые следы. Мы можем их увидеть, потому что разрешающая способность молекулярного метода исследования теперь фантастическая – один нуклеотид. Еще 15–20 лет назад это казалось чудом, волшебством. Геномы изучали с помощью скрещиваний мутантов, составляли генные карты, разрешение такого метода было не выше частей хромосом или групп генов, но уж никак не одного нуклеотида. Это все равно что заменить 20-кратную лупу на электронный микроскоп. И если уж эволюционные механизмы работают с наследственным материалом – последовательностями ДНК и РНК, то при такой разрешающей способности молекулярного метода мы это увидим.
Следы работы эволюционных механизмов бывают двух типов. Каждая аминокислота кодируется или одним, или, чаще, несколькими определенными тройками (триплетами) нуклеотидов. Если аминокислота кодируется одним-единственным триплетом, то замена любого нуклеотида в триплете приведет к замене аминокислоты; если она кодируется несколькими триплетами, то замена одного нуклеотида может привести либо к замене аминокислоты, либо к замене триплета другим из возможного набора, а аминокислота останется той же. Замена нуклеотида без замены аминокислоты называется синонимичной, а если аминокислота меняется, то получим несинонимичную, или значимую, замену. Если, сравнивая какой-то ген у двух организмов, мы видим, что среди наблюдаемых различий преобладают значимые замены, это указывает на действие положительного отбора, который поддерживал полезные мутации в этом гене. Если, наоборот, резко преобладают синонимичные замены, а значимых нет или очень мало, – это след отрицательного отбора, который отбраковывал все значимые мутации, потому что они оказывались вредными. Промежуточное соотношение указывает на нейтральную эволюцию. Таким образом, отношение синонимичных и несинонимичных замен свидетельствует о направлении отбора, и это первый тип «следов», оставляемых в геноме естественным отбором.
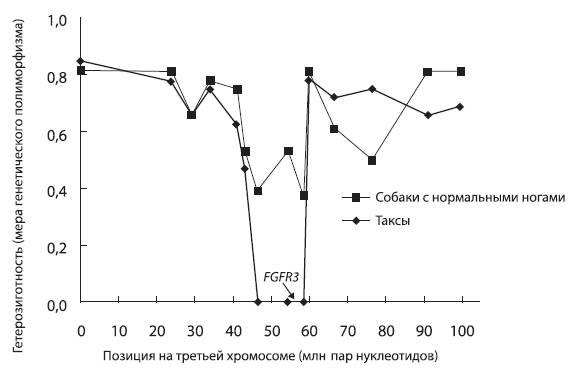
Пример «селективного выметания». У такс по сравнению с обычными собаками резко понижен генетический полиморфизм на участке третьей хромосомы, включающем ген FGFR3. Это указывает на то, что отбор, осуществлявшийся собаководами, выводившими новую породу коротконогих охотников на лис (было это менее 100 собачьих поколений назад), благоприятствовал какой-то мутации на этом участке хромосомы. В условиях отбора на «таксовость» некая мутация, произошедшая в этом участке, оказалась для собак «полезной», поскольку она обеспечивала успешное размножение, тогда как щенки, не имевшие этой мутации, отбраковывались селекционерами. Что же это за ген – FGFR3? Имеет ли он какое-то отношение именно к таксам? Оказывается, имеет. Белок, кодируемый этим геном, называется рецептором фактора роста фибробластов. Мутации в нем приводят к разнообразным нарушениям развития скелета, в том числе к укорочению конечностей. По рисунку из Pollinger et al., 2005.
Второй тип следов выявляется при изучении уровня полиморфизма (разнообразия) нуклеотидных последовательностей в популяции. Если мы отсеквенируем геномы множества представителей какого-нибудь вида и сравним их между собой, то увидим, что уровень полиморфизма, т. е. индивидуальных различий между геномами, распределен по геному неравномерно. Где-то полиморфизм выше – это те участки генома, в которых большинство мутаций оказываются нейтральными, поэтому полиморфизм свободно накапливается. Скорее всего, это просто не очень нужные участки, «генетический балласт». Где-то генетическое разнообразие ниже – это важные участки генома, в которых большинство мутаций оказываются вредными и удаляются очищающим отбором. Ну а в некоторых местах – и это самое интересное – мы увидим резкое, чуть не до нуля, снижение полиморфизма. В таких участках нет (или очень мало) не только значимых, но и синонимичных различий. Как правило, это означает, что здесь поработала «метла» положительного отбора. Это явление так и называют – selective sweep, т. е. «выметание посредством отбора». В середине участка с резко пониженным полиморфизмом обычно сидит какая-то полезная мутация. Она возникла не очень давно у какой-то особи, а потом быстро распространилась под действием положительного отбора. Вместе с мутацией автоматически распространялись и прилегающие к ней участки ДНК. Это явление называют сцепленным наследованием. До появления полезной мутации уровень полиморфизма в данном участке хромосомы был, скорее всего, примерно таким же, как на соседних участках. Но, когда отбор начинает распространять полезную мутацию, он автоматически распространяет и ее окружение со всеми индивидуальными (и в основном нейтральными) особенностями, присущими вовлеченному в сценарий участку ДНК. Повезет тем нейтральным аллелям, которые находились рядом с полезной мутацией. Остальные нейтральные вариации исчезнут из генофонда, а выживут те, что были у счастливого обладателя первого экземпляра мутантного гена. Как будто все варианты, кроме одного, из данного фрагмента генома «выметаются».
Со временем следы метлы стираются за счет накопления новых нейтральных мутаций. Таким образом, глубокие ямы на графике распределения полиморфизма указывают на относительно недавние случаи действия положительного отбора.
Как в первую неделю жизни хомячат работает мутантный сигнальный белок Agouti – пока неизвестно. Зато удалось показать, что данная мутация, по всей видимости, появилась и начала распространяться в популяции совсем недавно – позже, чем отступил ледник и сформировались Песчаные Холмы с их светлым грунтом. В пользу этого свидетельствуют результаты статистических тестов. В частности, оказалось, что «светлые» варианты гена Agouti (в которых отсутствуют вышеупомянутые три нуклеотида) меньше варьируют по остальным полиморфным сайтам, чем «темные» варианты. Уровень полиморфизма в окрестностях полезной мутации относительно низок. Это типичный пример «выметания посредством отбора», и этого не должно было бы наблюдаться, если бы данная мутация (выпадение трех нуклеотидов) существовала в популяции в качестве нейтральной задолго до того, как появились Песчаные Холмы и она стала полезной.