Полная версия
Эволюция. Классические идеи в свете новых открытий



Но это исключения. Как правило, эволюционное движение большого и сложного современного белка ограничено одним горным массивом – одной группой родственных функций. На больших высотах ландшафт приспособленности белков сильно разобщен, фрагментирован, так что с одного массива на другой попасть почти невозможно. Но вот на малых высотах, у подножья величественных горных массивов, простирается холмистая местность, передвигаться по которой простым и примитивным древним белкам было намного легче. Среди искусственно синтезированных коротких белковых молекул с произвольной последовательностью аминокислот можно найти молекулы, выполняющие – пусть и с низкой эффективностью – различные функции, выполняемые природными белками в клетке. Это значит, что, передвигаясь по этим низинам, можно наткнуться на предгорья какого-нибудь горного массива даже методом случайных блужданий!
Скорее всего, основные функции белков, соответствующие белковым надсемействам, были «нащупаны» еще в самом начале эволюционного становления генетического кода и синтеза белка у древних РНК-организмов[16]. Потом эти функции бесконечно совершенствовались и подразделялись на множество вариаций. Эволюционирующие последовательности взбирались все выше по склонам «своих» горных массивов, разбредаясь по развилкам и отрогам, и чем выше они поднимались, тем меньше оставалось у них шансов перейти с однажды выбранной горной системы на какую-то другую.
По-видимому, только для самых простых и коротких (но при этом все же полезных) белковых молекул существует реальная вероятность возникновения «из ничего», «на голом месте» – из случайной комбинации нуклеотидов, случайно закодировавших некоторую последовательность аминокислот. Такой способ формирования новых белков исчерпал себя еще в РНК-мире. С тех пор новые белки появляются только из старых – не с нуля, а путем модификации того, что есть. Это одно из главных правил эволюции. Почти все новое – это перекроенное старое.
Нейтральные мутации и генетический дрейф – движение без правил
Ландшафт приспособленности – образ яркий и полезный, но, как и всякая модель, он несовершенен. Многие аспекты эволюционного процесса с его помощью отразить трудно или невозможно. Реальный ландшафт приспособленности переменчив (впрочем, как и реальные горные системы). Если один белок в клетке изменился, это хоть немного, но изменит свойства клетки, ее поведение и внутреннюю среду – а значит, и «требования», предъявляемые отбором к другим белкам. Их ландшафты приспособленности станут немного другими. Изменение одного вида в сообществе неизбежно повлияет на факторы отбора, действующего на другие виды, и т. д.
Кроме того, трудно представить себе такой ландшафт, который точно отразил бы реальное соотношение вредных, полезных и нейтральных мутаций. Как правило, большинство мутаций нейтральны – по крайней мере у высших организмов, эукариот, у которых в геномах много участков, слабо влияющих на приспособленность. На втором месте по численности вредные мутации, на последнем – полезные. Нелегко вообразить склон такой формы, чтобы из каждой точки было больше разных путей, параллельных горизонту, чем путей, ведущих вверх или вниз. Но реальные склоны реальных ландшафтов приспособленности именно таковы.
Нейтральные мутации – это по определению такие мутации, которые не влияют на приспособленность, или, что то же самое, не подвергаются действию отбора. Нам пора познакомиться поближе с этим самым распространенным в природе классом мутаций. Забудем на время о ландшафте приспособленности и обратимся к другой модели, которая описывает процессы, происходящие с генетическими вариантами (аллелями) в популяции.
Допустим, у нас есть маленькая популяция мюмзиков из 40 особей. Для простоты примем, что мюмзики гаплоидны, т. е. имеют одинарный набор хромосом – один-единственнный экземпляр генома, а не два, как у нас с вами, диплоидных организмов. Достигнув возраста в один год, каждый мюмзик рожает нескольких детенышей, а потом сразу умирает от старости. Все детеныши одинаковые, но выжить из них могут только 40 – больше мюмзиков просто не помещается в старой железной бочке на дне пруда, где живет изучаемая популяция. Смертность детенышей случайна – ровно 40 случайно выбранных везунчиков станут взрослыми и через год произведут потомство.
Поскольку смертность случайна, приспособленность мюмзиков определяется только их плодовитостью, т. е. числом детенышей. Плодовитость зависит от генотипа. Допустим, у мюмзика есть один ген, влияющий на плодовитость. Обозначим его буквой А. Между прочим, не смейтесь: такая модель вполне годится для изучения некоторых законов популяционной генетики.
Некогда у всех мюмзиков был только один вариант (аллель) гена А. Обозначим его А1. Но потом у одной особи произошла мутация, и в результате появился второй вариант гена – аллель А2. Допустим (опять-таки для простоты), что в нашей популяции, когда мы приступили к ее изучению, у половины мюмзиков ген А был представлен первым вариантом, у другой половины – вторым. Таким образом, частота аллеля А2 равна 0,5 (q2 = 0,5) и частота аллеля А1 такая же (q1 = 0,5).
Нам нужно ответить на вопрос: как будет меняться со временем частота аллеля A2, если мутация, которая привела к его возникновению, была нейтральной?
Раз мутация была нейтральной, значит, плодовитость обладателей обоих аллелей одинакова. Допустим, они все рожают ровно по десять детенышей. Разумеется, потомство наследует родительский аллель гена А.
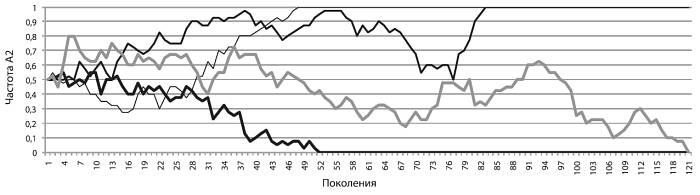
Может показаться, что раз аллели имеют одинаковую приспособленность, то их так и останется поровну. Это неверный ответ. На рисунке(см. ниже) показан результат четырех запусков нашей модели. Мы видим, что частота А2 во всех четырех случаях хаотически колебалась (по-научному такие колебания как раз и называются случайными блужданиями). Блуждания продолжаются до тех пор, пока частота аллеля не упрется либо в верхнюю «точку невозврата» (q2 = 1, частота аллеля достигла 100 %, аллель зафиксировался в генофонде), либо в нижнюю (q2 = 0, частота упала до нуля, аллель элиминировался из генофонда).
Так бывает всегда. Если аллель нейтрален, его частота будет «случайно блуждать» между нулем и единицей до тех пор, пока не упрется либо в верхний, либо в нижний предел. Рано или поздно нейтральный аллель либо зафиксируется (достигнет частоты 1), либо элиминируется – исчезнет из генофонда. Третьего не дано. Таким образом, блуждания хоть и случайны, их исход предсказуем. Мы точно знаем, что дело кончится либо фиксацией, либо элиминацией. Чем больше популяция, тем в среднем дольше придется ждать исхода[17], но он все равно неизбежен.
Здесь самое время сказать, что случайные, не зависящие от приспособленности изменения частот аллелей называются генетическим дрейфом. Под властью дрейфа находятся все нейтральные мутации (а в маленьких популяциях отчасти также и вредные с полезными, но об этом позже).
Можно ли вычислить вероятность того, что нейтральный аллель в конце концов зафиксируется, а не элиминируется? Да, это просто. В нашем примере аллели А1 и А2 исходно находились в равном положении, потому что у обоих была частота 0,5. Очевидно, что в такой ситуации шансы на фиксацию у них должны быть одинаковыми и равными 0,5. В половине случаев А1 зафиксируется, А2 элиминируется, в половине – наоборот.
Ну а если конкурирующих нейтральных аллелей не два, а, скажем, четыре и начальные частоты у них тоже одинаковые (0,25)? В этом случае дело кончится фиксацией одного из аллелей и элиминацией трех остальных, причем для каждого аллеля вероятность фиксации равна 0,25. Таким образом, очевидно, что вероятность фиксации нейтральной мутации в будущем просто-напросто равна ее частоте в данный момент: Pfix = q.
Если вы, дорогие читатели, еще не устали от этой примитивной математики, то позвольте познакомить вас еще с двумя простыми, интересными и полезными формулами.
Сколько нейтральных мутаций будет фиксироваться в популяции в каждом поколении? (Имеются в виду мутации уже не в одном и том же, а в разных генах.) Если мы сумеем это вычислить, то получим прекраснейший инструмент – молекулярные часы. Тогда мы сможем по количеству нейтральных генетических различий определить, когда жил последний общий предок сравниваемых организмов.
Вывод этой формулы – подлинный шедевр «биологической математики». Судите сами. Определим сначала, сколько новых мутаций появляется в популяции в каждом поколении. Будем считать для простоты, что подавляющее большинство мутаций нейтральны (это недалеко от истины). Ответ очевиден: U × N, где U – темп мутагенеза (среднее число новых мутаций у каждой новорожденной особи), N – численность популяции. Определить U можно, просто сравнивая геномы детей и родителей.
Теперь нужно понять, какая часть из этих U × N только что появившихся мутаций в итоге зафиксируется. Это и будет искомая величина – темп фиксации нейтральных мутаций в генофонде популяции за одно поколение. Обозначим ее буквой V.
Мы уже знаем, что вероятность фиксации мутации равна ее частоте: Pfix = q. Чему же равна частота только что появившейся мутации? Это совсем просто. Поскольку мутация только что появилась, она пока есть только у одной особи. Следовательно, ее частота равна 1/N. Вот, собственно, и все. Умножаем количество новых мутаций на вероятность фиксации каждой из них (т. е. на долю мутаций, которые в итоге зафиксируются) и получаем ответ: V = U × N × 1/N. Поразительно! Численность популяции волшебным образом сокращается, и величина N уходит из уравнения. Мы приходим к выводу, что темп фиксации нейтральных мутаций не зависит от численности и равен просто-напросто скорости мутагенеза: V = U. Не знаю, как вы, дорогие читатели, а мы, биологи, приходим в восторг от такой красоты.
Нам остался один шаг до молекулярных часов. После того как два вида, произошедшие от общего предка, разделились, в их генофондах независимо накапливаются нейтральные мутации. За время t (измеряемое в поколениях) первый вид накопит V × t = U × t нейтральных мутаций, и второй вид накопит столько же. Совместными усилиями они накопят 2U × t нейтральных отличий друг от друга. Зная темп мутагенеза U и подсчитав число различий между геномами сравниваемых видов (обозначим его буквой D), определяем время жизни их последнего общего предка: t = D/2U. Это и есть знаменитые молекулярные часы.
Если бы темп мутагенеза и скорость смены поколений были одинаковыми у всех живых существ, все было бы совсем просто. Но они, конечно, неодинаковы, поэтому в вычисления приходится вносить поправки. А еще есть проблема насыщения: в какой-то момент – обычно по прошествии десятков, а чаще сотен миллионов лет – гены разделившихся когда-то видов оказываются настолько «переполнены» нейтральными различиями, что величина D перестает расти, хотя нейтральные мутации продолжают фиксироваться. Ниже мы увидим пример исследования, показавшего, как уровень сходства между «случайно блуждающими» последовательностями приблизился к минимально возможному и дальше снижаться уже не мог.
К счастью, разные участки генома накапливают нейтральные изменения с очень разной скоростью[18]. Быстро меняющиеся участки ДНК используются для датировки недавних событий, а те, что меняются медленно, хороши для датировки событий глубокой древности.
Дрейф и отбор: кто кого?
Генетический дрейф царствует над нейтральными мутациями (аллелями), отбор – над полезными и вредными. Отбор, повышающий частоту полезных мутаций, называют положительным. Отбор, отбраковывающий вредные мутации, – отрицательным, или очищающим.
Впрочем, все так хорошо и просто только в больших популяциях. В маленьких ситуация сложнее, потому что отбор и дрейф – две главные движущие силы эволюции – начинают конкурировать друг с другом за контроль над слабовредными и слабополезными мутациями.
Посмотрим, как это происходит. Воспользуемся для этого той же моделью с мюмзиками. Изменим только характер мутации, которая привела к появлению аллеля А2. До сих пор мы считали, что мутация была нейтральной. Поскольку она была нейтральной, обладатели аллелей А1 и А2 имели одинаковую плодовитость. Давайте теперь предположим, что мутация была полезной, что она повысила плодовитость мюмзиков на 5 %. Это можно смоделировать так: пусть мюмзики с генотипом А1 рожают по 20 детенышей, а мюмзики с генотипом А2 – по 21. Начальная частота аллеля А2 пусть будет по-прежнему равна 0,5. Только теперь мы рассмотрим популяции с разной численностью (N).
На рисунке (см. ниже) показано, как будет меняться частота А2 в том случае, если его носители имеют пятипроцентное адаптивное преимущество. Мы видим, что в большой популяции (N = 5000) частота А2 неуклонно растет, приближаясь к единице. Так работает отбор в идеальных для него условиях, т. е. в больших популяциях, где влияние дрейфа на полезные и вредные аллели пренебрежимо мало. Форма у кривой довольно правильная, и это наводит на мысль, что ее можно описать какой-нибудь математической формулой. Это действительно так, но выводить формулу мы не будем, чтобы не утомить читателей (а любители математики могут сделать это самостоятельно)[19].
Перед нами, между прочим, важнейший эволюционный процесс – аллельное замещение, т. е. вытеснение более приспособленным аллелем менее приспособленного. Процесс идет не слишком быстро. В большой популяции, например состоящей из миллиона особей, для того чтобы зафиксировалась новая полезная мутация, дающая 5-процентное адаптивное преимущество, требуется около 560 поколений. А ведь 5 % – это серьезное преимущество. Такие мутации – редкость. Ждать, пока зафиксируется мутация с преимуществом в 1 %, придется уже 2800 поколений! Тем не менее в большой популяции отбор «чувствует» даже самую незначительную разницу в приспособленности. Это обеспечивает эффективную (пусть и медленную) фиксацию слабополезных мутаций и элиминацию слабовредных[20].
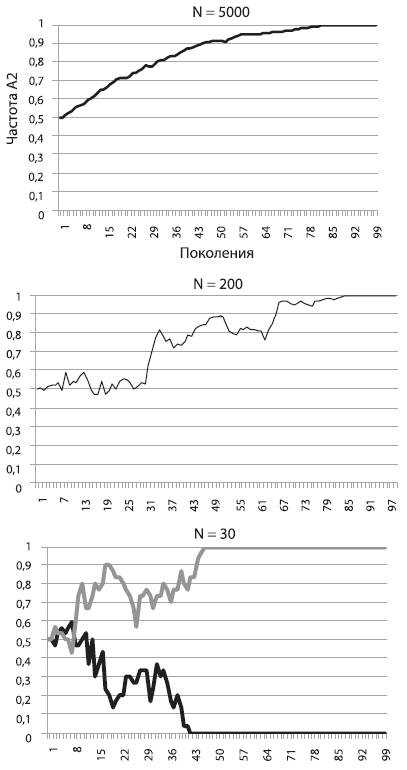
На среднем графике мы видим, какая судьба ждет такую же полезную мутацию, повышающую приспособленность на 5 %, в популяции меньшего размера (N = 200). Частота полезного аллеля и в этом случае росла и в конце концов достигла 1 (мутация зафиксировалась), но путь был труден и извилист. Пожалуй, нашей мутации могло и не повезти, ее шансы на фиксацию не были стопроцентными.
Наконец, на правом графике мы видим, что происходит с точно таким же полезным аллелем в совсем крошечной популяции (N = 30). На рисунке показаны результаты двух запусков модели. В одном случае мутация зафиксировалась, в другом – элиминировалась. Не правда ли, картинка похожа на результат работы дрейфа, а не отбора?
Так оно и есть. В этом заключается главный урок, который мы можем извлечь из наших экспериментов. Чем меньше популяция, тем слабее в ней власть отбора и тем могущественнее дрейф. В маленьких популяциях слабополезные и слабовредные мутации начинают вести себя фактически как нейтральные. Их частоты «случайно блуждают», пока не наткнутся на верхний или нижний порог. В маленьких популяциях, где царствует дрейф, небольшие различия в приспособленности становятся невидимыми для отбора. Поэтому слабовредная мутация запросто может зафиксироваться, а слабополезная – элиминироваться.
Хорошо это или плохо? В большинстве случаев, конечно, плохо. Потеря слабополезных мутаций мешает маленькой популяции приспосабливаться к меняющимся условиям. Бесконтрольное накопление слабовредных мутаций и вовсе может поставить ее на грань вымирания. Именно в этом, между прочим, видят биологи одну из причин того, что крупные животные вымирают в среднем чаще, чем мелкие. У крупных животных, таких как носороги или слоны, популяции не могут быть такими же большими, как у мышей или насекомых. Это снижает приспособляемость крупных животных.
Но у этой медали есть и обратная сторона. У маленьких популяций больше шансов выйти из «ловушки локального оптимума», т. е. сползти с невысокого пика ландшафта приспособленности и взобраться на другой, повыше. Ведь отбор гонит организмы вверх и только вверх. Если популяция велика и малейшее различие в приспособленности «заметно» для отбора, спуск по склонам становится невозможен. Однажды взобравшись на одинокий холм, большая популяция уже никогда с него не слезет. Что касается дрейфа, то он ведет организмы по ландшафту приспособленности хаотическим образом, не замечая подъемов и спусков. Если популяция невелика и дрейф силен, у организмов есть шанс иногда двигаться не только вверх, но и немного вниз (и в сторону). Спустившись в ложбинку, организмы могут «обнаружить», что отсюда есть другой, более перспективный подъем. Если, конечно, не вымрут раньше, чем на него наткнутся.
Вселенная древних белков продолжает расширяться
В 2010 году журнал Nature опубликовал интересную статью об эволюционном движении белков по ландшафтам приспособленности (Povolotskaya, Kondrashov, 2010). Авторы работы решили сравнить аминокислотные последовательности 572 древних белков, которые имелись уже у последнего общего предка всего живого[21] и были унаследованы его потомками, а также нуклеотидные последовательности соответствующих генов 836 прокариот (бактерий и архей). Эти белки называют древними, потому что после более чем 3,5 млрд лет эволюции в телах разнообразных потомков LUCA они до сих пор сохранили сходство своих аминокислотных последовательностей (вплоть до поразительного 40-процентного сходства рибосомных белков L14 у бактерий и архей) и продолжают выполнять те же функции, что и у Луки.
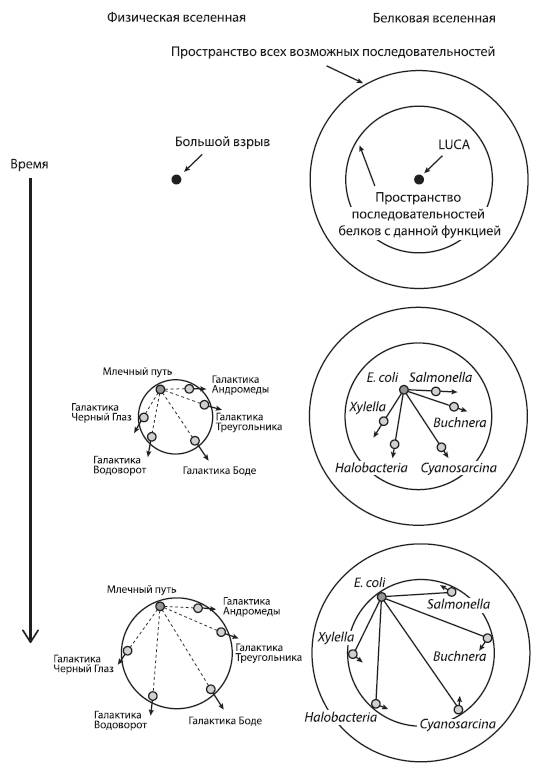
Расширение физической и белковой Вселенной. Стрела времени направлена сверху вниз. Слева: в ходе расширения физической Вселенной увеличиваются расстояния от произвольно выбранного объекта (например, Земли) до других объектов, причем скорость удаления пропорциональна расстоянию. Справа: в ходе расширения «белковой вселенной» дочерние молекулы удаляются в пространстве последовательностей от исходной точки, которая соответствует одному из белков LUCA. При этом дистанции между произвольно выбранным объектом (например, белком, принадлежащим эволюционной линии, которая привела к кишечной палочке E. coli) и другими объектами (родственными белками других эволюционных линий) постепенно растут. Однако белки с данной функцией не могут выйти за пределы «своей» подобласти в пространстве последовательностей – иначе это будет уже другой белок, выполняющий другую функцию (эволюционные события, связанные со сменой функции белка, в обсуждаемой работе не рассматриваются). Границы этой функциональной области показаны внутренним кругом. Рано или поздно расходящиеся белки достигнут этой границы, и дальнейшее расхождение станет невозможным. Стрелочками показано направление движения белков в пространстве последовательностей. По рисунку из Povolotskaya, Kondrashov, 2010.
Для начала Поволоцкая и Кондрашов решили выяснить, достигли уже эти белки максимального расхождения в пространстве последовательностей или их расхождение (накопление различий) все еще продолжается. Сохранение сходства после 3,5 млрд лет независимой эволюции, казалось бы, говорит о том, что возможности изменения этих белков ограниченны. Можно предположить, что области в пространстве последовательностей, соответствующие их функциям, невелики и предел расхождения давно достигнут. Но возможно и другое объяснение. Эти области могут быть велики, но труднопроходимы. Например, они могут представлять собой лабиринт из узких хребтов, разделенных пропастями, и поэтому на «освоение» всего доступного пространства не хватило миллиардов лет, прошедших со времен LUCA.
Исследователи проводят аналогию между эволюцией белков и расширением Вселенной. Эдвин Хаббл обнаружил, что галактики удаляются друг от друга, причем расстояние между галактиками положительно коррелирует со скоростью их разлетания. Экстраполируя эту тенденцию в прошлое, Хаббл пришел к выводу, что разлетание должно было начаться из одной точки. Эта идея легла в основу теории Большого взрыва. Нечто подобное происходит и с белками, расходящимися от общего предка – исходного белка с данной функцией, который был закодирован в геноме LUCA.
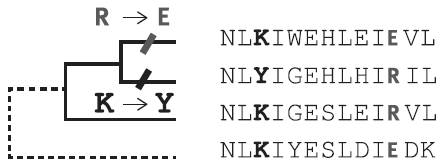
Принцип анализа белковых последовательностей, примененный Поволоцкой и Кондрашовым. Использовались четверки гомологичных белков, родственные отношения между которыми изображены в виде дерева. Анализ такой четверки позволяет понять, растет или снижается сходство между первыми двумя белками и четвертым. В данном случае считается, что у первого белка «предковая» (имеющаяся у белков 2 и 3) аминокислота R в 11-й позиции (выделена жирным шрифтом) заменилась на E, что привело к росту сходства первого белка с четвертым, т. е. к их сближению в пространстве последовательностей. У второго белка «предковая» аминокислота K в 3-й позиции (выделена полужирным шрифтом) заменилась на Y, что привело к снижению сходства второго белка с четвертым, т. е. к их расхождению в пространстве последовательностей. По рисунку из Povolotskaya, Kondrashov, 2010.
Чтобы выяснить, закончилось ли уже расширение «вселенной древних белков» или оно продолжается до сих пор, авторы применили оригинальные методы анализа белковых последовательностей. Нам будет полезно с ними ознакомиться – хотя бы для того, чтобы понять логику биоинформатики, молодой науки, занимающейся сравнением и осмыслением генетических текстов.
Использовались четверки гомологичных (происходящих от одного предка) и выполняющих одну функцию белков. Первые два белка в каждой четверке – это близкородственные молекулы с похожими аминокислотными последовательностями. Эти два белка назывались «сестринскими», и именно их эволюция анализировалась в рамках данной четверки.
Третий белок отличался от сестринских белков сильнее, чем они друг от друга. Он выполнял роль «внешней группы», позволяя понять, какие аминокислотные замены произошли в первом, а какие во втором сестринском белке. Например, если у внешнего белка в данной позиции стоит аминокислота K (лизин) и такая же аминокислота стоит здесь у первого из двух сестринских белков, а у второго в этом месте находится другая аминокислота (например, Y – тирозин), то считалось, что K в данной позиции – это «предковое», исходное состояние, а у второго сестринского белка произошла замена K на Y.
К этим трем белкам добавлялся четвертый, еще более далекий от сестринских, чем третий. Если у четвертого белка в данной позиции стоит аминокислота K (как на рисунке), то делался вывод, что у второго сестринского белка произошло эволюционное изменение, увеличившее дистанцию между белками 2 и 4: белки разошлись. Если бы у четвертого белка здесь стояла аминокислота Y, то был бы сделан вывод, что в результате изменения, затронувшего белок 2, белки 2 и 4 сблизились.
В общей сложности было обработано 13,6 млн таких четверок. Для каждой четверки определялось число замен, ведущих к сближению последовательностей и к их расхождению. По соотношению этих двух величин можно понять общую тенденцию: расходятся ли белки, сближаются или балансируют вокруг некого постоянного уровня сходства последовательностей.
Оказалось, что даже у наиболее удаленных друг от друга гомологичных белков тенденция к расхождению значительно преобладает над тенденцией к сближению. Следовательно, «вселенная древних белков» продолжает расширяться и пределы областей, соответствующих их функциям, за 3,5 млрд лет так и не были достигнуты. Эволюция этих белков была крайне медленной. Что же ее сдерживало? Решению этой проблемы посвящена вторая часть статьи, приводящая на память другой рассказ Борхеса – «Сад расходящихся тропок».
Лабиринт расходящихся тропок
Обычного отрицательного отбора, отсеивающего вредные мутации, явно недостаточно для того, чтобы объяснить медленную эволюцию древних белков. Предположение о том, что каждая аминокислота, стоящая в данной позиции, влияет на приспособленность одинаковым образом независимо от «контекста» (т. е. от того, какие аминокислоты стоят в других позициях в том же белке или в других белках, взаимодействующих с ним), соответствует представлению о легкопроходимом ландшафте приспособленности (см. рисунок ниже). Но со времен LUCA в каждом синонимичном сайте (так называют те нуклеотиды в цепи ДНК, изменение которых не влияет на структуру кодируемого белка) произошло уже свыше 100 замен. Синонимичные нуклеотидные замены – это пример движения по ровным горизонтальным поверхностям ландшафта приспособленности. Из этого следует, что в случае хорошей проходимости ландшафта изучаемые белки давным-давно должны были освоить всю потенциально доступную им область в пространстве последовательностей, и тогда мы не наблюдали бы их продолжающегося расхождения.