Полная версия
Вирус, который сломал планету. Почему SARS-CoV-2 такой особенный и что нам с ним делать
Самого по себе связывания с ACE2 уже достаточно для проникновения. Разумеется, не каждая вирусная частица, зацепившаяся за рецептор, сумеет попасть внутрь клетки, однако при существенном количестве вируса довольно у многих это получится. Но коварные вирусы придумали[6] дополнительный механизм, радикально увеличивающий шансы на проникновение: они используют сидящие в мембране протеазы[7] – ферменты, расщепляющие белки. Такие протеазы чрезвычайно важны для клетки, так как очень многие белки изначально синтезируются в виде полуфабрикатов – длинных аминокислотных цепей. Для того чтобы перевести такие заготовки в рабочую форму, их нужно разрезать в одном или нескольких местах – иногда чтобы откусить лишний хвостик, иногда потому, что свернуться в молекулы правильной формы могут только короткие цепочки. Множество таких протеаз находится в мембране большой внутриклеточной фабрики всевозможных полезных веществ под названием аппарат Гольджи. Но некоторые выносятся на внешнюю мембрану, вероятно для того, чтобы завершить процесс производства белков, которые активны на поверхности клетки или выделяются вовне, например, разнообразных сигнальных молекул. Еще часть протеаз оказывается на внешней мембране как побочный продукт внутриклеточной логистики – ферменты попадают туда в пузырьках с разными экспортными белками, регулярно поставляемыми из аппарата Гольджи.
Протеазе все равно что расщеплять, лишь бы у белка была определенная последовательность, которую она узнаёт и режет. Хитрый коронавирус обзавелся сразу несколькими такими последовательностями в своем спайк-белке. После того как SARS-CoV-2 связывается с клеточным рецептором, он «подставляет» эти последовательности протеазам – и те простодушно расщепляют S-белок в нужных местах. Как предполагается, после этого конформация (форма) образовавшихся кусочков изменяется таким образом, что вся вирусная частица подтаскивается максимально близко к поверхности клетки и коронавирусу остается только слить свою мембрану с клеточной – точно так же, как сливаются вместе две капли масла, плавающие на воде. По другой гипотезе, вирус попадает внутрь клетки путем эндоцитоза, то есть как бы проваливается в выпячивающийся внутрь клетки мембранный пузырек{5}. Но как бы то ни было, содержимое вирусной частицы оказывается внутри клетки.
Использовать клеточные протеазы для проникновения внутрь умеют многие вирусы: например, злобный родственник нынешнего коронавируса SARS (вирус, вызвавший вспышку атипичной пневмонии в 2002–2004 годах) эксплуатировал протеазу под названием TMPRSS2. Но SARS-CoV-2 не ограничился одной протеазой. В его спайк-белке вдобавок к последовательности, узнаваемой TMPRSS2, есть фрагмент, который расщепляет клеточная протеаза фурин. Предполагается, что готовность «работать» сразу с двумя протеазами делает нынешний коронавирус намного более заразным, чем его предшественник: если почему-либо TMPRSS2 окажется недоступна, он всегда сможет воспользоваться альтернативной протеазой{6}. Более того, благодаря использованию протеаз SARS-CoV-2 скрывается от иммунной системы. Важнейшая часть вирусной оболочки, на которую реагируют различные иммунные компоненты, в том числе антитела, – RBD-фрагмент. До того как какая-нибудь из протеаз расщепит спайк-белок, этот кусочек находится в «лежачей» конформации и практически не выдается наружу. После взаимодействия с протеазой RBD-фрагмент поднимается над поверхностью и его можно легко обнаружить, но в этот момент вирус уже сливается с клеточной мембраной, и, для того чтобы его обезвредить, необходимо привлекать другие рода иммунных войск{7}.
НЕВОЛЬНЫЙ ПОМОЩНИКРазличные протеазы используют в своих целях многие вирусы. Фурин печально знаменит тем, что сотрудничает с особо неприятными из них. Сайт (специфический участок) для расщепления фурином есть, например, у высокопатогенных штаммов птичьего гриппа. Его несет находящийся на поверхности вирусной частицы белок гемагглютинин. После того как сидящий на внешней мембране клетки фурин разрежет гемагглютинин на две субъединицы, на одной из них формируется участок, облегчающий слипание вирусной и клеточной мембран. Еще раз вирус птичьего гриппа использует фурин для того, чтобы более эффективно выпускать наружу новосинтезированные вирусные частицы – в этом случае работа пептидазы облегчает слияние вирусной и клеточной мембран изнутри клетки. Помогает фурин и вирусу иммунодефицита человека (ВИЧ): один из белков его оболочки синтезируется в виде полуфабриката и нуждается в разрезании фурином. Белки флавивирусов, к которым относятся возбудители таких опасных болезней, как энцефалит, желтая лихорадка или лихорадка денге, также расщепляются фурином в процессе сборки вирусных частиц.
Удобную протеазу научились использовать не только вирусы: многие бактерии с ее помощью активируют свои токсины. Фурин расщепляет на две субъединицы А и В токсин дифтерийной палочки, после чего субъединица А отправляется в ядро и тормозит процессы, необходимые для синтеза клеточных белков (сам синтез белков идет в цитоплазме, но для его регуляции необходима работа определенных ядерных генов). После того как фурин разрежет на три части токсин сибирской язвы, структурная часть получившихся кусочков формирует канал, через который ядовитые фрагменты проникают в цитоплазму{8}.
Казалось бы, такой вредный для клетки белок должен исчезнуть под давлением отбора, но увы, фурин и другие протеазы играют важнейшую роль в работе клеток и развитии эмбрионов, поэтому животным приходится носить в себе такую мину. По этой же причине – из-за участия фурина во множестве физиологических процессов – его вряд ли получится использовать как мишень для потенциального лекарства от COVID-19, хотя в экспериментах на культурах клеток и показано, что его блокировка уменьшает инфекционность SARS-CoV-2. Предыдущие попытки применить ингибиторы фурина (вещества, которые «выключают» его) для лечения других заболеваний показали, что такой подход дает множество побочных эффектов{9}{10}.
Размножение
Попасть внутрь клетки – половина дела. Цель вируса – создать как можно больше собственных копий, которые смогут распространиться и заразить другие клетки. Для этого необходимо синтезировать тысячи новых молекул РНК{11} для загрузки в вирусные частицы, а также все необходимые белки. Своих ресурсов для этого у коронавируса нет, зато есть инструменты, при помощи которых он может заставить клетку выполнить требуемые задачи. Эти инструменты – особые белки, которые переключают клетку из нормального режима работы в режим пособничества вирусу. Информация об аминокислотной последовательности таких хакерских белков закодирована в вирусной геномной молекуле РНК, причем это сделано крайне изобретательно. Чтобы впихнуть все необходимые данные в относительно небольшой геном, коронавирус (и не он один) использует хитрую комбинаторику. Его гены не записаны в молекуле РНК один за другим: они расположены внахлест – то есть перекрываются. Благодаря такому сжатию в одной и той же РНК умещается информация о большем количестве белков, чем если бы гены шли подряд. Если вы готовы немного погрузиться в биологию, чтобы разобраться, как именно вирусы извлекают информацию из перекрывающихся генов, читайте врезку ниже. Если нет – пропустите ее и переходите к следующему абзацу.
СДВИНУТЬ И РАЗРЕЗАТЬЗаложенную в молекуле РНК информацию считывает клеточная молекулярная машина рибосома. Она выглядит как округлая коробочка с длинной щелью, сквозь которую протягивается нить РНК. В активной рабочей зоне щели в каждый момент времени находятся три нуклеотида – буквы генетического кода. Как вы помните из главы 1, каждая такая тройка кодирует одну аминокислоту – базовую единицу белка. Вокруг рибосомы в цитоплазме плавают все 20 аминокислот, которые переносит специальный транспорт – особым образом свернутые маленькие молекулы РНК. Их называют тРНК, и каждой аминокислоте соответствует строго определенная тРНК. Аминокислоты на своем транспорте могут заплывать в щель рибосомы. Если оказавшаяся там аминокислота «правильная», то есть кодируется именно той тройкой нуклеотидов, которые в этот момент сидят в центре щели, тРНК «прилипнет» к ним и рибосома присоединит переносимую этой тРНК аминокислоту к растущей белковой цепи. После этого рибосома сдвинется по молекуле РНК на три нуклеотида и процесс будет повторяться до момента, пока машинка по синтезу белка не наткнется на стоп-кодон – определенную тройку нуклеотидов, на которых она отвалится от РНК.
Для того чтобы рибосома могла синтезировать с одной РНК несколько белков, вирусы используют хитрый трюк. Недалеко от стоп-кодона последовательность их РНК устроена таким образом, что легко закручивается в петлю. Рибосома умеет расплетать такие структуры, но на это ей требуется время. Петля только часть ловушки. Непосредственно перед ней находится особый участок РНК, который называют скользким. Он состоит из идущих подряд одинаковых букв, и, когда рибосома разбирается с петлей, она может случайно сдвинуться на одну букву, не заметив этого. Раскрутив петлю, рибосома продолжит синтез белка, но его последовательность окажется измененной, так как все следующие тройки нуклеотидов тоже будут сдвинуты на одну букву. В случае SARS-CoV-2 именно так синтезируются белковые цепи ORF1a и ORF1b (см. рис. 3). ORF1a рибосома строит как положено – от начала цепи до стоп-кодона. ORF1b получается, когда она запинается на расположенной ближе к концу ORF1a петле, перескакивает на одну букву и доделывает цепь до стоп-кодона в конце ORF1b. Таким образом, после нескольких циклов синтеза в клетке оказываются более короткие цепи ORF1a и длинная цепь ORF1ab, которая почти целиком включает ORF1a – кроме нескольких последних нуклеотидов, на которых рибосома соскользнула, – и всю цепь ORF1b.
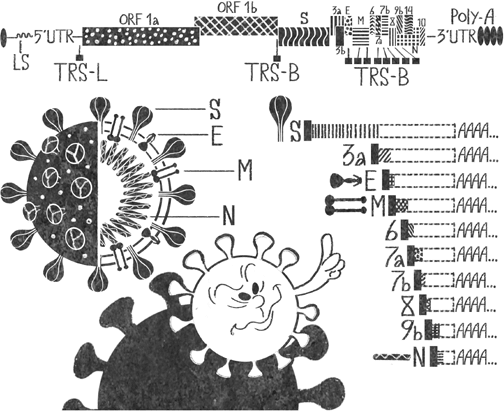
Рис. 3. Благодаря различным приемам сжатия контента в относительно небольшом геноме SARS-CoV-2 закодировано очень много информации. Попав в клетку, вирус первым делом подставляет рибосомам левую часть своей РНК. Они синтезируют с нее две длинные полипептидные цепочки ORF1a и ORF1b, причем часть последовательности ORF1b находится внутри ORF1a, но сдвинута на несколько нуклеотидов. Оставшиеся белки синтезируются с другого конца геномной РНК (так называемый 3’-конец) по одному – то есть для каждого из них РНК-зависимая РНК-полимераза сначала создает отдельную мРНК, на основе которой рибосома строит белок. Полимераза начинает считывать информацию с 3’-конца, доходит до одной из последовательностей TRS-B, пробуксовывает на них и либо сразу перепрыгивает на участок TRS-L, где заканчивает синтез, либо считывает информацию дальше – до следующего участка TRS-B.
UTR – untranslated region, нетранслируемые области, «сервисные» участки РНК. Белки их не синтезируются, но эти регионы необходимы для регуляции процесса.
PolyA – поли(А)-хвост, участок РНК, составленный только из аденина. Необходим для регуляции жизненного цикла молекулы РНК
Сначала с вирусной РНК клеточными рибосомами синтезируются две длинные белковые цепи ORF1a и ORF1ab (см. рис. 3), причем вторая половина длинной цепи оказывается сдвинута относительно первой из-за того, что рибосома поскальзывается на особой последовательности и продолжает синтез уже с нового места. Эти цепи разрезаются на отдельные белки, которые выполняют две задачи: 1) обеспечить бесперебойный синтез всех компонентов вирусных частиц и 2) не дать клетке-хозяину сообщить иммунной системе, что произошло заражение. Разрезает ORF1a и ORF1ab находящаяся в них же вирусная протеаза. После того как зараженная клетка порабощена, синтезированный с вирусной РНК фермент со сложным названием РНК-зависимая РНК-полимераза считывает со второй половины той же РНК оставшиеся гены – это в основном гены структурных белков, необходимых для сборки вирусных частиц. И уже на основе образовавшихся коротких молекул РНК рибосомы строят вирусные белки. Синтезировать белки непосредственно с исходной вирусной РНК нельзя: их последовательности как бы накладываются друг на друга (здесь вирус тоже использует сжатие информации), поэтому ген каждого белка нужно получить в виде отдельной молекулы РНК. Сложное объяснение – во врезке, его опять же можно пропустить.
ГЕНОМНЫЕ ПРЫЖКИВ случае с синтезом коротких РНК SARS-CoV-2 механизм получения множества молекул с одной матрицы иной. РНК-зависимая РНК-полимераза начинает считывать информацию с «правого» конца большой вирусной РНК (см. рис. 3). В какой-то момент она доходит до особой последовательности TRS (transcription-regulating sequence – последовательность, регулирующая транскрипцию, то есть синтез РНК) и пробуксовывает на ней из-за особой структуры TRS. Дальше полимераза либо продолжает считывать информацию до следующей TRS, либо перепрыгивает сразу на «левый» концевой участок, где особый сигнал заставляет ее прекратить работу. Прыжок или продолжение – вероятностные процессы, и после нескольких циклов в цитоплазме нарабатываются РНК всех генов второй половины исходной РНК.
Полный список белков коронавируса SARS-CoV-2 и их функции описаны в таблице 1. Аббревиатурой nsp обозначают неструктурные белки (от английского non-structural proteins). К ним относятся сервисные белки вируса вроде РНК-зависимой РНК-полимеразы и белков, обеспечивающих сборку вирусных частиц, а также nsp, выключающие собственный метаболизм клетки и мешающие ей позвать на помощь иммунную систему. Остальные белки составляют каркас вирусной частицы.

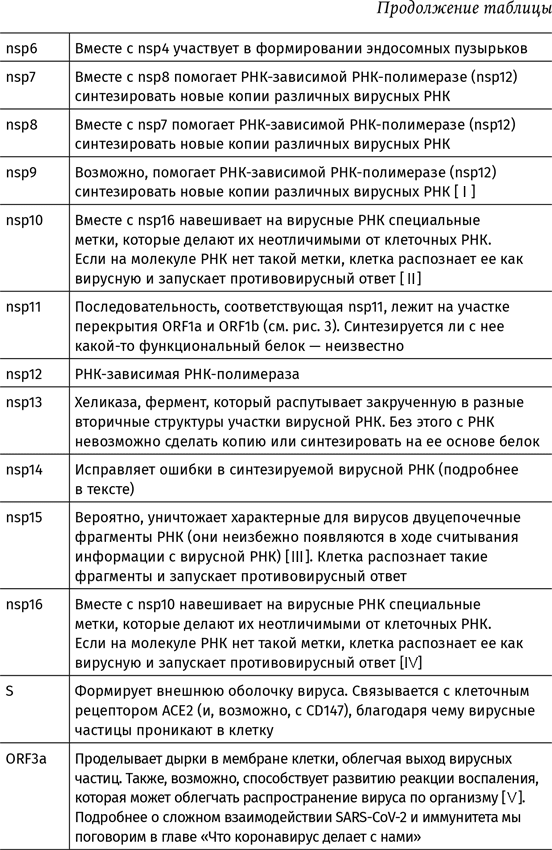
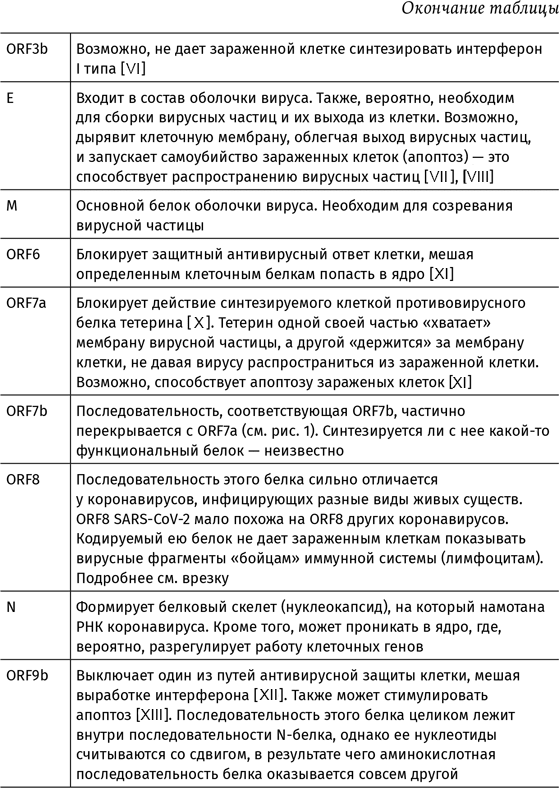
В геноме SARS-CoV-2 видны еще три последовательности, с которых, теоретически, могли бы считываться белки – ORF9c, ORF10 и ORF14. Но действительно ли они «работают», или это просто артефакт – неизвестно.
[I] M.-P. Egloff et al., “The severe acute respiratory syndrome-coronavirus replicative protein nsp9 is a single-stranded RNA-binding subunit unique in the RNA virus world,” Proc. Natl. Acad. Sci., vol. 101, no. 11, pp. 3792–3796, Mar. 2004.
[II] Y. Wang et al., “Coronavirus nsp10/nsp16 Methyltransferase Can Be Targeted by nsp10-Derived Peptide In Vitro and In Vivo To Reduce Replication and Pathogenesis,” J. Virol., vol. 89, no. 16, pp. 8416 LP-8427, Aug. 2015.
[III] Там же.
[IV] V. D. Menachery, K. Debbink, and R. S. Baric, “Coronavirus non-structural protein 16: Evasion, attenuation, and possible treatments,” Virus Res., vol. 194, pp. 191–199, Dec. 2014.
[V] K. Siu et al., “Severe acute respiratory syndrome Coronavirus ORF3a protein activates the NLRP3 inflammasome by promoting TRAF3-dependent ubiquitination of ASC,” FASEB J., vol. 33, no. 8, pp. 8865–8877, Aug. 2019.
[VI] Y. Konno et al., “SARS-CoV-2 ORF3b is a potent interferon antagonist whose activity is further increased by a naturally occurring elongation variant,” bioRxiv, p. 2020.05.11.088179, Jan. 2020.
[VII] D. Schoeman and B. C. Fielding, “Coronavirus envelope protein: current knowledge,” Virol. J., vol. 16, no. 1, p. 69, Dec. 2019.
[VIII] “UniProtKB – P59637 (VEMP_CVHSA). Envelope small membrane protein,” UniProt. [Online]. Available: https://www.uniprot.org/uniprot/P59637. [Accessed: 14-Aug-2020].
[XI] M. Frieman, B. Yount, M. Heise, S. A. Kopecky-Bromberg, P. Palese, and R. S. Baric, “Severe Acute Respiratory Syndrome Coronavirus ORF6 Antagonizes STAT1 Function by Sequestering Nuclear Import Factors on the Rough Endoplasmic Reticulum/Golgi Membrane,” J. Virol., vol. 81, no. 18, pp. 9812–9824, Sep. 2007.
[X] J. K. Taylor et al., “Severe Acute Respiratory Syndrome Coronavirus ORF7a Inhibits Bone Marrow Stromal Antigen 2 Virion Tethering through a Novel Mechanism of Glycosylation Interference,” J. Virol., vol. 89, no. 23, pp. 11820–11833, Dec. 2015.
[XI] S. R. Schaecher and A. Pekosz, “SARS Coronavirus Accessory Gene Expression and Function,” in Molecular Biology of the SARS-Coronavirus, Berlin, Heidelberg: Springer Berlin Heidelberg, 2010, pp. 153–166.
[XII] C.-S. Shi et al., “SARS-Coronavirus Open Reading Frame-9b Suppresses Innate Immunity by Targeting Mitochondria and the MAVS/TRAF3/TRAF6 Signalosome,” J. Immunol., vol. 193, no. 6, pp. 3080–3089, Sep. 2014.
[XIII] C.-S. Shi, N. R. Nabar, N.-N. Huang, and J. H. Kehrl, “SARS-Coronavirus Open Reading Frame-8b triggers intracellular stress pathways and activates NLRP3 inflammasomes,” Cell Death Discov., vol. 5, no. 1, p. 101, Dec. 2019.
Глядя на таблицу 1, можно подумать, что мы уже очень много знаем о белках SARS-CoV-2. На самом деле это не так. Большинство представленных здесь сведений получены учеными, исследовавшими вирус SARS. Геномные последовательности SARS и SARS-CoV-2 сходны на 79 %{12}, и большинство белков практически неотличимы друг от друга. Но очевидно, с каждым месяцем исследователи будут лучше разбираться в подрывной деятельности нового коронавируса. Так, предварительный анализ взаимодействия белков SARS-CoV-2 с белками клетки показал, что даже безобидные структурные белки теоретически могут вмешиваться в те или иные иммунные процессы{13}. Чем лучше мы будем понимать, как именно коронавирус воздействует на организм, тем больше у нас будет потенциальных слабых мест, на которые можно нацеливать лекарства.
Об одном белке нового коронавируса нужно упомянуть отдельно. Это nsp14 – фермент, умеющий исправлять ошибки, допущенные при копировании вирусных РНК. В первой главе мы говорили, что РНК-содержащие вирусы отличаются повышенной склонностью мутировать. Одна из причин – помимо нестабильности самой молекулы РНК – это крайне неряшливая работа РНК-зависимой РНК-полимеразы, фермента, который синтезирует новые копии вирусной РНК. По сравнению, например, с ДНК-полимеразами, которые копируют молекулы ДНК, она допускает колоссально много ошибок[8]. Так много, что это угрожает выживанию вируса: если не исправлять их, очень быстро последовательность изменится настолько, что на ее основе нельзя будет синтезировать ни одного работающего белка. Поэтому коронавирусы обзавелись ферментом-пруфридером (от английского proofreading, то есть правка текста корректором, в ходе которой он исправляет ошибки). Он «видит» неправильно вставленные РНК-полимеразой нуклеотиды и вырезает их. После этого полимераза вновь пытается вставить в это место нуклеотид – и так как ошибается она, все же, не каждый раз, со второй попытки опечатка часто исчезает. Впервые исправляющий ошибки белок обнаружили у SARS, и, похоже, это уникальная придумка именно коронавирусов. По крайней мере, ни у каких других РНК-вирусов такого фермента не найдено.
МОЛЕКУЛЯРНЫЙ ДИВЕРСАНТУспех SARS-CoV-2 во многом связан с его способностью обманывать клеточные противовирусные механизмы. Этим занимаются минимум девять вирусных белков (см. табл. 1). Часть из них мешают клетке синтезировать интерфероны, часть маскируют вирусные РНК так, чтобы они стали неотличимы от клеточных, часть уничтожают следы деятельности вируса (двуцепочечные РНК), которые выдают его специальным белкам-«охранникам». Некоторые из этих механизмов были у SARS и MERS, но один является абсолютным эксклюзивом нынешнего коронавируса: последовательность ORF8 кодирует белок, который не дает клетке выставлять на поверхности фрагменты собственных белков{14}. Такая самопрезентация – чрезвычайно важный механизм, позволяющий иммунной системе вовремя заметить и уничтожить подозрительные клетки, неважно, завелся в них вирус или они мутировали и готовятся стать раковыми. Каждая клетка нашего организма – как и организмов всех позвоночных животных – в обязательном порядке отчитывается перед клетками иммунной системы с неприятным названием Т-киллеры (или цитотоксические Т-лимфоциты). Для этого клетка разрезает небольшой процент всех белков, которые обнаружит у себя в цитоплазме, на небольшие кусочки и выставляет их на поверхность на особых белках MHC I (major histocompatibility complex class I, белки главного комплекса гистосовместимости класса I). Т-киллеры регулярно наведываются к MHC I, и, если какой-нибудь из выложенных фрагментов покажется им неправильным (то есть совпадет с чужеродным белковым мотивом, на который настроен каждый Т-киллер), клетку немедленно убивают. ORF8 мешает клетке производить MHC I, направляя свежесинтезированные белки главного комплекса гистосовместимости класса I в лизосомы – заполненные «едкими» ферментами пузырьки, в которых клетка уничтожает всякий мусор. Уменьшая количество MHC I, SARS-CoV-2 минимизирует шансы, что какой-нибудь из них подцепит и выставит на обозрение Т-киллерам фрагмент вирусного белка.
Похожими трюками балуются и другие неприятные вирусы, например ВИЧ и аденовирус. И хотя клетки с подозрительно малым количеством MHC I на поверхности тоже уничтожаются иммунной системой, но, пока она заметит, что белков не хватает, пройдет достаточно много времени и вирус успеет как следует размножиться. Отчасти с этим связана высокая патогенность SARS-CoV-2: раз попав в организм, он с ходу обманывает иммунную систему и выигрывает критические первые часы, пока вирусных частиц еще мало и иммунитет мог бы задавить инфекцию в зародыше.
Выход из клетки
Новые вирусные частицы собираются не просто в цитоплазме: SARS-CoV-2 организует фабрику по их производству в особом отделе клетки – эндоплазматическом ретикулуме. Это огромный лабиринт, стенки которого сделаны из той же мембраны, что и клеточная оболочка. В эндоплазматическом ретикулуме клетка синтезирует часть белков и снабжает их особыми метками-«пропусками», направляющими эти белки в разные клеточные «департаменты». В том числе упакованные в мембранные пузырьки экспортные белки отправляются во внешнюю среду. Ферменты вируса приостанавливают синтез собственных клеточных белков и оккупируют эндоплазматический ретикулум. Свежесинтезированные структурные белки SARS-CoV-2 встраиваются в мембранные пузырьки, внутрь загружается намотанная на N-белок вирусная РНК – и новая вирусная частица готова. Множество вирусов набиваются в большой мембранный пузырек – такой же, в который в норме клетка упаковывает свои «экспортные» белки, – и плывут к наружной мембране. Пузырек сливается с ней, и паразиты выходят в межклеточное пространство искать новых жертв. Жизненный цикл коронавируса повторяется.
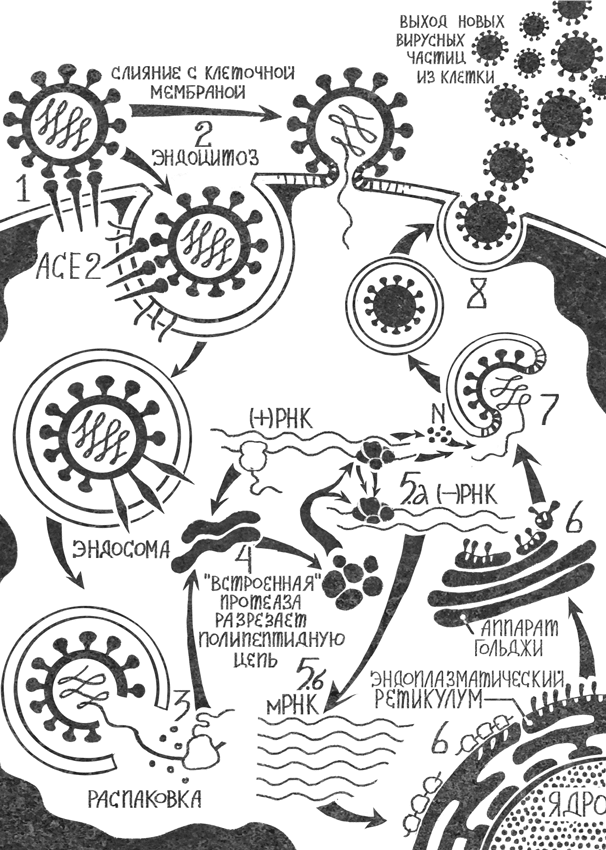
Рис. 4. После того как спайк-белок вируса связывается с рецепторами ACE2 (1), клеточная протеаза фурин или TMPRSS2 разрезает спайк (не показано на рисунке) и вся частица подтягивается ближе к поверхности клетки. Изначально предполагалось, что после этого ее мембрана сливается с клеточной ((2), верхняя стрелка), но позже появились данные, что вирус попадает внутрь клетки путем эндоцитоза ((2), нижняя стрелка). Оказавшись в клетке, вирус выпускает наружу свою геномную РНК, которая выходит из эндосомного пузырька в цитоплазму, где клеточные рибосомы начинают синтезировать с нее длинную полипептидную цепь (3). Один из входящих в нее белков – протеаза, и она разрезает цепь на отдельные белки (4). В том числе протеаза высвобождает РНК-зависимую РНК-полимеразу, которая немедленно начинает синтезировать новые копии вирусной геномной РНК (5.а) и оставшиеся вирусные сервисные и структурные белки (5.б). Последние отправляются в эндоплазматический ретикулум, где происходит сборка новых вирусных частиц (6). Она продолжается в аппарате Гольджи, а затем в заготовки будущих вирусов загружается геномная РНК и уплотняется, наматываясь на пришедший из цитоплазмы новосинтезированный N-белок (7). В мембранных (эндосомных) пузырьках готовые частицы подплывают изнутри к клеточной мембране, пузырьки сливаются с ней, и новые вирусные частицы оказываются снаружи клетки (8)
Почему SARS-CoV-2 такой особенный
Описанные выше трюки коронавируса выглядят весьма хитроумными, но в действительности примерно так ведут себя очень многие вирусы. За миллиарды лет гонки вооружений со своими хозяевами они идеально отточили навыки захвата и порабощения чужих клеток[9]. И все же у некоторых вирусов это получается лучше остальных. Например, из семи известных коронавирусов, которые способны заражать человека, только три – SARS, MERS и нынешний SARS-CoV-2 – представляют серьезную опасность, остальные же вызывают банальные простуды. SARS убил 10 % всех заразившихся, MERS – 34 %, от SARS-CoV-2 умирает, видимо, около 1 % инфицированных (точнее подсчитать можно будет только после того, как закончится активная фаза эпидемии). При этом пандемию устроил самый безобидный из «суровых» коронавирусов – SARS-CoV-2, – потому что научился отлично передаваться от человека к человеку. SARS и тем более MERS делали это существенно хуже.