


Полная версия
Sea-Birds
The family Alcidae (the auks) take the place in the north of the penguins of the south. Undoubtedly their origin has been in or not far from the Bering Sea. Of the twenty-two species, sixteen belong to the northern part of the North Pacific, twelve to the Arctic Ocean north of the Circle, and six to the northern part of the North Atlantic.
This concludes the list of sea-birds belonging to groups of super-family or higher status whose evolution has been marine. There are several further (secondarily marine) groups which contain sea-birds, or part-time sea-birds; thus all four members of the order Gaviae the divers, breed in the Arctic, and North Atlantic and Pacific regions, and winter at sea on the coasts of the oceans. Many of the twenty species of grebes, order Podicipedes, are marine outside the breeding-season, and six of them visit the coasts of the North Atlantic at that time. Among the geese and ducks many (see Appendix, see here) are partly marine, and some (e.g. eiders and scoters) are largely marine in the breeding—as well as in the off-season: two eiders and three scoters breed in the North Atlantic-Arctic. Among the waders (Charadriidae) the subfamily Phalaropinae contains only three members, all of which breed in the Arctic, North Atlantic and North Pacific, and two of which winter in the open sea.
If we ignore these secondary sea-birds, and consider the 267 species of the primary marine groups, we find that the hierarchy is this: South Pacific 128 (51 per cent.); North Pacific 107 (40 per cent.); North Atlantic 74 (28 per cent.); South Atlantic 73 (27 per cent.); Indian Ocean 73 (27 per cent.); Antarctic 44 (16½ per cent.); Arctic 31 (11½ per cent.); Mediterranean 24 (9 per cent.); and purely inland only 7 (2½ per cent.).
It can be seen that the North Atlantic, with its seventy-four species, is much lower than either half of the Pacific than would appear warranted by its area. There is not the faintest hint, from the radiation of any of the sea-bird groups, that either North or South Atlantic has been the arena of any great evolutionary changes. The Atlantic has been colonised from without; by penguins from the Antarctic; by petrels from the South Pacific; by pelecaniform birds and terns probably from the Indian Ocean; by gulls and auks from the Arctic. The North Atlantic and the immediately neighbouring parts of the Arctic have but two present sea-bird genera and only thirteen species of their own. We need not be surprised at this indication that the Atlantic’s bird fauna is derived from that of other oceans if we accept Wegener’s theory of the origin of the Atlantic; but whether the Wegener theory is true or not it is quite clear that the North Atlantic has not been the home in which any important group of sea-birds has evolved. This is not to say that there has been no sea-bird evolution in the North Atlantic; but it has not usually gone beyond the differentiation of species. Of this it has, indeed, much to show. Some of the classic examples which E. Mayr (1942) has discussed are North Atlantic species. Mayr’s thesis is that one species can only become two after it has been differentiated geographically. He opposes the notion which has found favour in some quarters that speciation may occur by ecological differentiation or by the differentiation of behaviour.
So far the available evidence appears to uphold Mayr’s view—at all events, for birds. During the present century much systematic work in the description and measurement of birds has been conducted in American and European museums, and much practical and theoretical work on evolution has also been done. But it needed the persuasions of Mayr and Julian Huxley (1942), amongst a few others, to collate the work of the systematists and the evolutionary zoologists. Sea-birds lend themselves to evolutionary study because they are so largely confined to coasts for breeding purposes. This makes their distribution often linear rather than of the ordinarily spatial two-dimensional type; and this linear distribution makes it easy to apply Huxley’s concept that the characteristics of animals tend to grade from one part of their range to another in an orderly way. Some of these gradations had been recognised long before Huxley thought of the word “cline” because they are adaptations to the environment. For instance Bergmann’s Rule states that from the warmer parts of an animal’s distribution-area to the colder parts there tends to be an increase in its size. Thus the puffins, black guillemots and eider-ducks of the Arctic are considerably bigger than those of Britain. The main adaptive reason for this is that larger animals have less surface in proportion to their weight, and consequently heat is not lost from them (if warm-blooded) so rapidly as it is from small animals. Another rule, Allen’s Rule, states that warm-blooded animals of cold climates tend to have their heat-radiating surfaces decreased by a reduction in size of their extremities and limbs such as ears, tails, necks, legs and noses. There is also a general tendency (Gloger’s Rule) for animals to become darker as humidity increases.
If we examine those sea-birds which are widely distributed, we find clines in various characteristics, notably in size, i.e. total size, and also size of limbs and extremities, beak-length, wing-length, etc., and in colour. There are also clines in shape; for instance the fulmars of the north-east Atlantic have very thick bills, those of Baffin Island rather more slender bills, those of the North Pacific more slender bills still, and those of the Antarctic very slender bills indeed. No sea-bird is arranged quite evenly in its geographical distribution. Just as the distribution in space is never even, so are the gradations in character never even. From one part of the geographical distribution of a species to the other, change often occurs more as a series of steps rather than as continuous ramp.
Most working ornithologists today will agree that there are more subspecific names about than a true understanding of bird evolution requires. It is the species which has reality and significance. In this book we have tried to be sparing in the use of subspecies, and have rejected some that appear in many current text-books. Nevertheless, a study of the geographical races of the species of the North Atlantic sea-birds will lead us to examine here some of the more fascinating examples of geographic differentiation. The classic example among the sea birds is the chain of the Larus argentatus and fuscus group, the herring-gulls and lesser blackbacks, which may include some birds which are regarded as separate species, e.g. the California gull L. californicus, and the so-called ‘Iceland’ gull, or better the Greenland herring-gull, L. glaucoides or leucopterus. The relationships of this superspecies (Fig. 4a) were first worked out by B. Stegmann (1934): we have included the results of subsequent systematic work in this map (Fig. 4b) and in the discussion which follows.
It will be seen that the subspecies is composed of three chains of subspecies which unite in Central Siberia, where the resident breeding subspecies is Birula’s herring-gull Larus argentatus birulai. The two northerly chains link round the Polar Basin, the two end links of one overlapping with the two end links of the other. Where they overlap, the two races of one chain-end are ‘herring-gulls,’ of the other ‘lesser blackbacks.’ These behave as different species. It can be found convenient to make the ‘species’ separation in the chain, between the two races birulai and heuglini, thus calling the latter Larus fuscus heuglini (it is the first really dark-mantled gull in the chain). This is more practical than splitting the chain into argentatus and fuscus in the Bering Strait area, though this is probably the place of origin of the ancestral gull that gave rise to the whole chain; for if all the palearctic group were fuscus some confusion would surround the light-mantled Mediterranean forms.
Special comments can be made on various members of the chain. In the zone of overlap in Western Europe the herring-gulls are distinguished from the lesser blackbacks not only by form but by many habits. The lesser blackbacks breed often inland on moors, and when coastal tend to colonise flattish ground set back from the cliff-tops beloved of the herring-gulls. While the herring-gulls are dispersive in winter, the lesser blackbacks are almost entirely migratory, wintering south of all but their most southerly breeding-places, though some of the dark L. f. fuscus of Scandinavia winter in Britain, and recently a minority of the British race L. f. graellsii has ‘revived’ an old habit of wintering in England, especially in Cheshire and Lancashire. Both species are also extending their breeding-range north; L. a. argentatus has colonised east and north-east Iceland since 1909, and a herring-gull of this or the Scandinavian race omissus was breeding on Bear Island in 1932, though not 1948. The graellsii lesser blackback has established itself in south Iceland since about 1925, and a group intermediate between graellsii and fuscus in Denmark since 1922.
The North American situation is of great interest. As the herring-gulls range north-east they become generally paler in colour. The much-discussed Kumlien’s herring-gull L. a. kumlieni was for a long time held to be a hybrid between the ‘Iceland’ gull of Greenland and L. a. thayeri, Thayer’s gull of the Canadian Arctic and Thule corner of north-west Greenland. But there seems no doubt that it is a valid race (Taverner, 1933) with its own discrete breeding-distribution in southern Baffin Island, though on the western marches of its distribution there are apparently some forms intermediate between it and thayeri (Hørring, 1937) and colonies off south-west Baffin Island have been described as mixed (Soper, 1928).
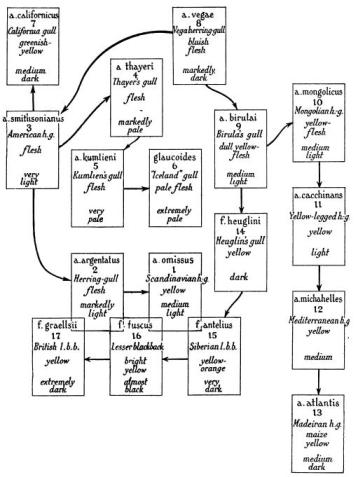
FIG. 4
Breeding distribution and relationships of all subspecies of Larus fuscus, L. argentatus and related forms. a Diagram of forms, with leg and mantle-colour
The palest of all the herring-gulls is the ‘Iceland’ gull. Unquestionably this extremely pale bird, with pale flesh legs, is a herring-gull, and conspecific with the other herring-gulls of North America. Reports of its breeding in the Canadian arctic archipelago are due to confusion with thayeri; there is no evidence whatever of its overlapping with this or any other subspecies of L. argentatus anywhere; and its similarity in size, structure and plumage is obvious. It is just a very pale kind of herring-gull; and at the same time happens, through convergence, to be extraordinarily similar to, though smaller than, the glaucous gull.* It is entirely confined to Greenland, breeding north to Melville Bay on the west (this inhospitable coast separates it from thayeri) and to Kangerdlugssuaq (at the south end of the Blosseville coast) on the east. Evidence of its breeding farther north in east Greenland, and elsewhere (e.g. Franz Josef Land, Novaya Zemlya) is quite unsatisfactory, and probably due to confusion with the glaucous gull; on Jan Mayen it was stated by F. Fischer to be as abundant as the glaucous gull in 1882–83, and to be nesting on low ledges, but it has not been proved to breed there since.
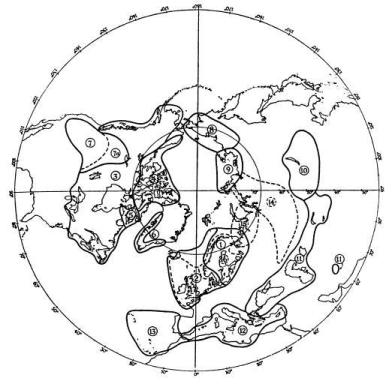
FIG. 4bBroken line: in Eurasia, L. fuscus; in North America southern limit of possible area of overlap between L. californicus and L. argentatus smithsonianus. Fig. 4a gives the key to the numbered forms.
In a complex situation, such as this, a confusion of scientific names is to be expected. In other cases it is often found that the vernacular name is less equivocal, and certainly more stable, than the scientific name! Such is not the present case, however; for the name ‘Iceland gull’ makes confusion worse confounded. It has never bred in Iceland. Hørring and Salomonsen (1941) have already used the English name Greenland Gull to describe it, and regard it as a race of Larus argentatus. We commend to our readers, and to the compilers of the Lists of the American and British Ornithologists’ Union : the Greenland Herring-Gull, Larus argentatus glaucoides (= L. a. leucopterus).
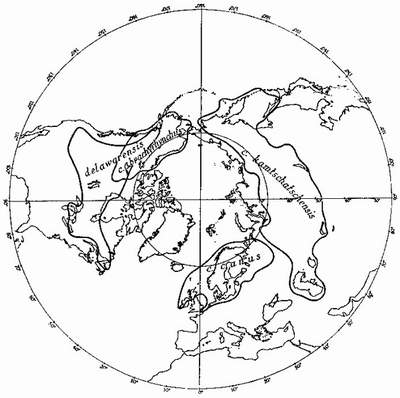
FIG. 5
Breeding distribution of Larus canus, the common gull, and the closely related L. delawarensis, the ring-billed gull
Among the North Atlantic sea-birds are others whose species have differentiated geographically and whose range-end populations have become different enough to occupy the same geographical area—but separate ecological niches, and thus preserve their identity. For instance, it is probable that the ring-billed gull Larus delawarensis, of North America, and the common gull of the Old World, L. canus, have not long since shared a common ancestor, though a subspecies of the common gull, which has probably spread across the Bering Straits from the Old World, now occupies Alaska and parts of the Canadian North-West, where it overlaps with the western element of the ring-billed gull (Fig. 5). Here the two act as different species. The glaucous gull and the great blackback, which overlap in eastern North America, Iceland and parts of the European Arctic (Fig. 6) may be not long ago descended from a common ancestor. They very rarely hybridise. How the three species of terns—the arctic, common and Forster’s—which are very closely related, arrived at their present distribution (Fig. 7) is difficult to imagine at this stage of their evolution, but they all may be descended from a common tern of north-east Asia or an arctic tern of the North Pacific—from which part of the world the species has probably spread, differentiated and overlapped.
Various suggestions could be made as to the origins of the two guillemots, the common and Brünnich’s guillemot (Fig. 8). Possibly the original guillemot was a common guillemot (Uria aalge) type which got divided into two subspecies in the Atlantic and Pacific by the Ice Age, but not before it had had time to give rise to an arctic race adapted to the harder life. After the Ice Age, with the ameliorating conditions, perhaps both the Atlantic and the Pacific guillemots began pushing north again, this time to meet and overlap with their arctic descendant, which, meantime, had differentiated sufficiently to offer no direct competition. It is interesting to note that the most arctic of the common guillemot races, Uria aalge hyperborea of Iceland, Novaya Zemlya, and Lapland, has a very thick bill and a considerable resemblance to Brünnich’s guillemot, with which it, however, does not interbreed, nor apparently compete. Perhaps it is recapitulating some of the early stages in the origin of Brünnich’s guillemot. To some extent Brünnich’s guillemot, with its razorbill-like beak, appears to replace the razorbill in the arctic, where it may occupy the same ecological (feeding and breeding) niche in relation to the common guillemot as the razorbill does in relation to that bird in the south part of the common guillemot’s range.
The student of variation will find much material for his researches among the North Atlantic sea-birds. Several species of North Atlantic birds, notably the common guillemot, the three smaller skuas and the fulmar, are polymorphic or dimorphic. They exist in several so-called phases. Some common guillemots have a white ring embracing their eye from which a white line runs back towards the back of their heads. These are called ‘bridled’ guillemots, and were for long actually thought to be of a different species. The phases of the skuas range from very light phases with yellow over their ears and the back of their necks, white throats and bellies, to those which are almost uniformly brown. The breeding fulmar population of Britain, the Faeroes, Iceland, Jan Mayen and West Greenland are all light-coloured with white bellies, necks and breasts, but in Baffin Island, Spitsbergen and Franz Josef Land the fulmars are nearly all very dark coloured. Between the light forms of Britain, etc., and the dark forms of Spitsbergen, there are a number of puzzling intermediates, most in evidence on Bear Island (and often to be seen at sea in the Rockall area), and the situation among the fulmars is therefore one not of dimorphism but of polymorphism, as it is among the skuas.
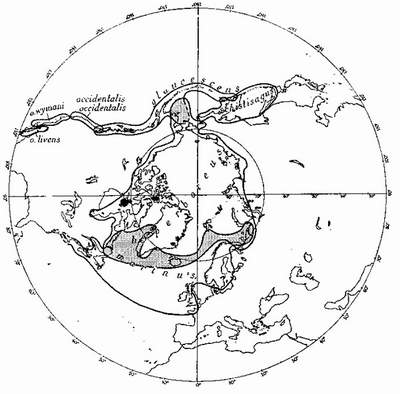
FIG. 6
Breeding distribution of a group of closely-related gulls: Larus occidentalis, the western gull: L. glaucescens, the glaucous-winged gull; L. schistisagus, the slaty-backed gull; L. hyperboreus, the glaucous gull; and L. marinus, the great black-back. Areas of overlap shaded. Black areas in Canadian Arctic represent outpost breeding-places of L. marinus
Southern, who has carefully studied the problem of the differential distribution of the bridled guillemot, thinks that its ‘bridle’ is probably controlled by a single Mendelian factor, which appears to control also a slight difference in the skull structure and the shape of the tail-feathers. He organised counts of the percentage of bridled guillemots throughout Britain in the years round 1939 and again in those round 1949; and he has also collected as much evidence as he could from the rest of the guillemot’s range. Two main conclusions are apparent: first, the percentage of bridled birds increases from SSE to NNW (with a reversal in Iceland); and secondly the percentage is not always constant at any one place—there are signs of trends towards increase or decrease, and of shifts, or drifts, of the balance. Possibly the possession of a bridle gives a guillemot an advantage over other guillemots in some environments, and a disadvantage in others, though we do not know why: the alternative is that possession of the bridle is the result of an advantageous mutation that is spreading through the population; which is unlikely to be the case on the evidence, though Southern has been careful to show that the possibility still exists. There is no indication that bridled guillemots prefer to mate with each other rather than with unbridled guillemots; mating in a mixed colony appears to be completely, or almost completely, at random.
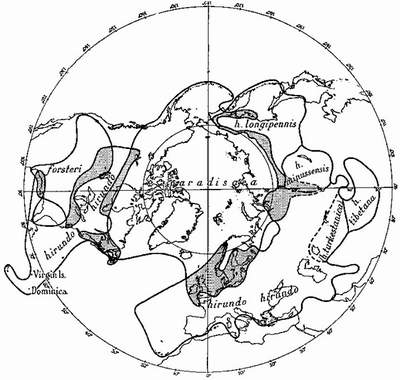
FIG. 7
Breeding distribution of three closely-related terns: Sterna hirundo, the common tern; S. paradisaea, the arctic tern; S. forsteri, Forster’s tern. Areas of overlap shaded. S. h. turkestanica is a doubtful subspecies
Southern shows that the percentage of bridled birds marches fairly closely with humidity and cloudiness; but, as he points out, many other factors may be involved. The changes between c.1939 and c.1949 may be linked with the climatic amelioration, but “might very well be due to random fluctuation.” The actual percentages as recorded in the paper of Southern and Reeve (1941) and Southern (1951), and in a few notes published by other observers, are shown on the maps (Figs. 9a, 9b). The results of Southern’s enquiry of 1949 have shown that out of the very many colonies studied in Britain at only five has a significant* change been recorded in ten years, four of which show decreases of the percentage of bridled birds and one an increase. One of the decreases is at St. Kilda, where the expedition of 1939 found 16.5 per cent. of the guillemots bridled and that of 1948 only 10.3 per cent. (one of us took part in both counts). Other decreases in Britain have been significant, as at the Isle of May, 5.3 to 3.2 in ten years; and at Unst in Shetland—23.8 to 16.9 per cent. in the same period. There has also been a significant decrease—of about one-third—in Iceland; thus at Grimsey in the north, from 8.7 in 1939 to 6.9 per cent. in 1949; at Hafnaberg, in south-west Iceland, from 29 per cent. in 1939 to 18.1 per cent. in 1949; in the Westmann Islands a parallel decline from 75 per cent. in 1935 (Lockley, 1936) to 50 per cent. in 1949.
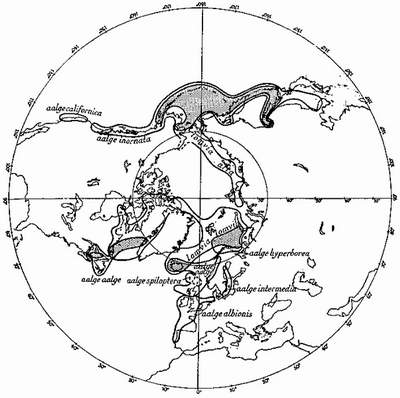
FIG. 8
Breeding distribution of two closely-related guillemots or murres: Uria aalge, the common guillemot; and U. lomvia, the arctic guillemot (Brünnich’s guillemot* in the Atlantic, Pallas’s murre in the Pacific). Areas of overlap shaded.
Increases noted in the 1939–1949 enquiries were several, but only one, at Foula in Shetland, was significant and by checked observers (from 24 per cent. in 1938 to 29.4 per cent. in 1948–49). Increases on the margin of significance were recorded from St. Bee’s Head in Cumberland, Marwick Head in Orkney, and the Fair Isle. Apart from these small increases in the last decade, there was a significant increase of the percentage on Noss in Shetland from 15.5 in 1890 to 26.5 in 1938, which seems great enough to embrace a possible slight observer-error.
Unfortunately, too few of the early bridled guillemot counts are reliable, though some from Berneray and Mingulay (‘Barra Head’) in the Outer Hebrides may be so. This had 20.2 per cent. in 1871; 12 in 1939; 9.8 in 1949; 12.6 in 1950. The decrease between 1871 and 1939 is significant, though the other apparent changes are not so. Elsewhere we have followed Southern in discarding such vague records as ‘about one in every nine or ten.’
Nothing is yet known about the percentage of bridled guillemots along the coast of Norway, except that it has remained slightly over 50 per cent., at Bear Island from 1932 to 1948. At the Karlov Islands off the Murmansk coast the percentage was 42 in 1938. It seems likely, from the rather scanty figures from Novaya Zemlya, which Southern slightly misdates and misplaces, that the percentage may be about the same on islets in Pukhovy and Bezymiannaya Bays off that island (36.4 and 50).
These changes are curious and it is clear that much remains to be solved about this interesting problem in distribution and evolution. Nor is much known about the distribution of the bridled form in the New World, save the following: H. F. Lewis found 128 bridled out of a sample of 724 (17.7 per cent.) in the colonies along Quebec Labrador in 1929. One of us found 51 bridled out of a sample of 295 (17.3 per cent.) at Cape St. Mary, on the south-west corner of the Avalon Peninsula of Newfoundland, in 1953. In June, 1940 at Funk Island and other parts of the east coast of Newfoundland within forty miles of it W. Templeman (1945) collected twelve common guillemots (? at random) of which six (50 per cent.) were bridled. When Hørring and Salomonsen (1941) compiled a list of all the common guillemots that had been then collected on the west coast of Greenland they recorded six out of thirty-two (18.7 per cent.) as bridled, but knew of no breeding-colony. Soon afterwards Salomonsen (1944) became aware of the colony in the Sukkertoppen district; but no count has apparently yet been made there.

FIG 9a The principal breeding-colonies of the common guillemot in Britain. The percentages of bridled forms in the breeding-populations, as determined chiefly by H. N. Southern and his colleagues, are shown. Minus and plus signs in brackets indicate changes in the decade c. 1939–c. 1949 which are significant, or on the borderline of significance. Crossed circles mark sites of former colonies.
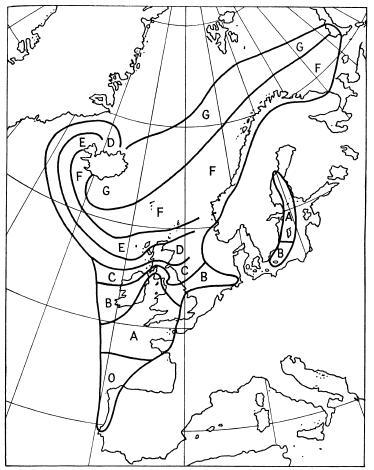
FIG. 9b The distribution of bridled guillemots in the East Atlantic breeding-populations: O: no bridled birds observed. A: under 1 per cent bridled. B: under 2 per cent bridled. C: under 5 per cent bridled. D: under 10 per cent bridled. E: under 20 per cent bridled. F: under 50 per cent bridled. G: over 50 per cent bridled.