


Полная версия
От атомов к древу: Введение в современную науку о жизни



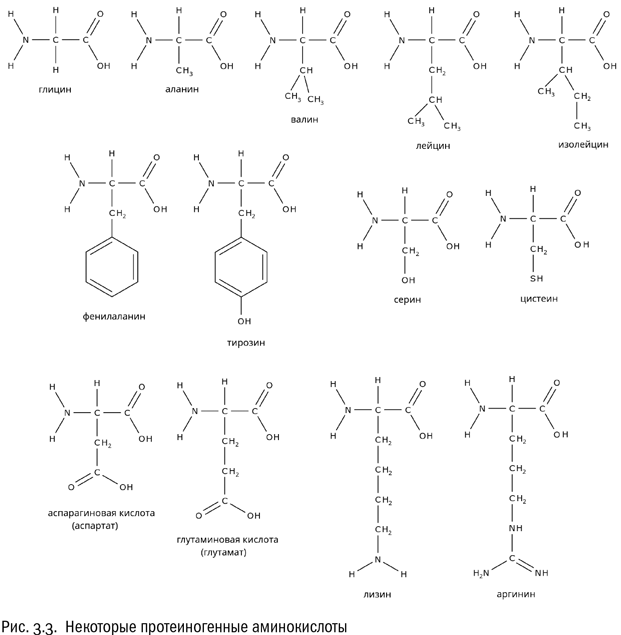
Есть аминокислоты, у которых в состав радикала входит гидроксильная группа – OH (серин, тирозин) или аналогичная ей, но содержащая вместо атома кислорода атом серы тиольная группа – SH (цистеин). В таких радикалах есть полярные связи, а потому они взаимодействуют с водой гораздо лучше. Эти радикалы – гидрофильные.
Все аминокислоты, перечисленные нами до сих пор, называются нейтральными. Это означает, что в водном растворе их молекулы электрически не заряжены. Мы уже видели, что в любой аминокислоте есть карбоксильная группа, свойства которой кислотные, и аминогруппа, свойства которой, наоборот, основные (см. главу 1). Попадая в воду, карбоксильная группа отдает протон и становится заряжена отрицательно (–COO–), а аминогруппа присоединяет протон и становится заряжена положительно (–NH3+). Суммарный заряд молекулы аминокислоты в результате остается равным нулю. Это – нейтральная молекула.
А что, если карбоксильных групп две? Тогда эта аминокислота будет в растворе заряжена отрицательно. И действительно, в состав белков входит пара таких аминокислот – это аспарагиновая и глутаминовая кислоты. У них обеих есть карбоксильная группа не только при альфа-атоме, но еще и в радикале. И, соответственно, этот радикал несет дополнительный отрицательный заряд.
Для краткости аспарагиновую и глутаминовую кислоты часто называют, соответственно, аспартатом и глутаматом. Тут надо пояснить одну тонкость, связанную с названиями веществ. Аспартат и глутамат – на самом деле названия не кислот, а их анионов или (что то же самое) их солей. Например, глутамат – это соль глутаминовой кислоты. В биохимии этим сплошь и рядом пренебрегают, используя названия кислот и солей как синонимы. Ведь что такое соль? Это кислота, у которой на месте протона оказался любой другой катион. Если же она диссоциирована и не имеет никаких катионов вообще (а это бывает в растворах очень часто), то за ней обычно ради удобства оставляют название соответствующей соли. Именно это мы на примере аспартата с глутаматом и видим. Название соли – это название аниона, в виде которого молекула реально существует в воде.
Глутамат (будем отныне называть его так) интересен не только тем, что участвует в образовании белков. В организмах подавляющего большинства животных он служит еще и нейротрансмиттером, причем одним из важнейших. В нервной системе человека глутамат используют для передачи возбуждения примерно 40 % нейронов – это очень много! Почти все основные информационные потоки в нашем мозге идут посредством выделения глутамата, служащего для нервных клеток возбуждающим сигналом. Выше мы упоминали, что нейротрансмиттером является и еще одна протеиногенная аминокислота, а именно глицин (см. главу 1). Но действие глицина тормозящее (то есть успокаивающее), а глутамата – именно возбуждающее. Поскольку глутамат входит в состав каких угодно белков, то его очень много в пище, но пищевой глутамат в мозг почти не попадает – нервные клетки синтезируют его сами.
Кроме того, к глутамату очень чувствительна наша вкусовая система. Обычно считается, что есть пять основных вкусов, для которых на языке существуют отдельные типы рецепторов: соленый, кислый, сладкий, горький и выделенный в начале XX века вкус умами. Соленый – это вкус поваренной соли, кислый – протонов (H+), сладкий – сахаров. Горький вкус – самый сложный, он не привязан к какому-то одному классу молекул и возникает как реакция на любое вещество из большой и разнообразной группы зачисленных мозгом в “ядовитые”, это эволюционно выработанный механизм защиты от токсичной пищи. Ну а умами – это не что иное, как вкус глутамата. Судя по всему, в ходе эволюции органов чувств позвоночных животных именно глутамат был выделен как индикатор вкуса белков (важнейших питательных веществ как-никак). Вот почему на языке для него есть особые рецепторы. Ощущение вкуса глутамата – это эволюционно выработанный сигнал о том, что в рот попало нечто белковое.
Глутамат часто добавляют в пищу, причем как в виде кислоты (пищевая добавка E620), так и в виде натриевой соли (пищевая добавка Е621). И раз уж мы заговорили об этом веществе, воспользуемся случаем, чтобы развеять несколько связанных с ним заблуждений. Может быть, кому-то пригодится.
Итак, во-первых, утверждение, что глутамат – усилитель вкуса, неточно. Выражение “усилитель вкуса” могло бы относиться к некоему (вымышленному) веществу без собственного вкуса и запаха, обладающему свойством обострять вкус любой еды. Глутамат этого не делает: у него просто есть свой вкус, точно так же, как у сахара или у соли. Механизмы восприятия вкуса глутамата и вкуса, допустим, того же сахара принципиально не отличаются друг от друга. Просто сахар воспринимается одними рецепторами, а глутамат – другими.
Во-вторых, неверно часто встречающееся мнение, будто “натуральный” глутамат (предположительно безобидный) – это совсем не то, что глутамат “искусственный” (предположительно вредный и опасный). Люди, которые так думают, просто не знают, о каком веществе идет речь. А мы с вами теперь знаем. Глутамат – это не какая-нибудь загадочная сложная смесь (состав которой действительно мог бы варьироваться), а одно-единственное химическое соединение, описываемое незатейливой формулой. Вот она, эта формула: HOOC–CH2–CH2–CH(NH2) – COOH. Только и всего. Глутамат, полученный искусственно, не отличается от полученного готовым из природных продуктов, потому что отличаться там нечему.
В-третьих, глутамат, получаемый с пищей, едва ли опасен для нервной системы прежде всего потому, что он в нее почти не проникает – это обеспечивается специальным физиологическим барьером. Нервные клетки синтезируют глутамат самостоятельно. К тому же в белковых продуктах (таких, как творог, мясо или соя) глутамата наверняка больше, чем попадает его в еду в качестве пищевой добавки, – просто потому, что это составная часть любых белков.
В-четвертых, на глутамат нет аллергии{27}. Аллергию вызывают чужеродные вещества, а не такие, которые жизненно необходимы и всегда присутствуют в организме в высоких концентрациях, – а глутамат как раз относится к последним.
Так что опасность глутамата – это, судя по всему, типичный современный миф.
Однако вернемся к другим аминокислотам. Если есть отрицательно заряженные аминокислоты, логично ожидать, что существуют и положительно заряженные. Это действительно так. Пример положительно заряженной аминокислоты – лизин, имеющий формулу NH2–CH2–CH2–CH2–CH2–CH(NH2) – COOH. Как видим, у лизина в радикале есть дополнительная аминогруппа, которая ведет себя так, как аминогруппе и положено: приобретает протон. Еще одна положительно заряженная аминокислота – аргинин, радикал которого включает довольно редкую (больше она нам нигде не встретится) гуанидиновую группу – NH – C(NH) – NH2, тоже охотно присоединяющую протон.
Таким образом, аминокислоты, из которых состоят белки, можно поделить на четыре категории: гидрофобные, гидрофильные нейтральные, положительно заряженные и отрицательно заряженные. Разумеется, все эти различия касаются только той части молекулы аминокислоты, которую мы назвали радикалом (R). “Базовая” часть (включающая атом углерода, атом водорода, карбоксильную группу и аминогруппу) во всех рассмотренных нами протеиногенных аминокислотах одна и та же. Особое положение занимает разве что самая простая из всех возможных аминокислот – глицин, у которого вместо радикала атом водорода.
Аминокислоты и связи
Итак, любой белок – это полипептид, то есть цепочка аминокислот, соединенных пептидными связями. На самом деле теоретически можно придумать полипептид, не являющийся белком, но мы сейчас поступим проще и будем считать, что эти слова – синонимы. В подавляющем большинстве тех случаев, которые нам могут встретиться, так оно и есть.
Для начала представим, что молекулу полипептида бросили в воду. Очевидно, что она не останется там вытянутой в прямую линию, а будет как-то сворачиваться. Это сворачивание будет зависеть от взаимодействия аминокислотных остатков как с молекулами воды, так и между собой. В целом пептидная цепь устроена довольно просто: ее “скелет” образуют пептидные группы, соединяющие между собой альфа-атомы углерода, а радикалы торчат в стороны. Все эти части огромной молекулы как-то размещаются в пространстве относительно друг друга, и в результате белок приобретает свою трехмерную форму – как обычно говорят, конформацию. Белок с нарушенной конформацией, как правило, совершенно бесполезен для организма. Поэтому соблюдение конформации – это очень важно.
Как же она складывается? Есть четыре типа взаимодействий между аминокислотами, определяющих объемную структуру белка, в который они входят.
Во-первых, это водородные связи (см. главу 2). В белке их обычно много, они возникают и между пептидными группами, и между боковыми цепями аминокислот (“боковая цепь” и “радикал” – в данном случае синонимы). Особенно это относится к тем аминокислотам, радикалы которых нейтральны и гидрофильны – вроде, например, серина или тирозина.
Во-вторых, это гидрофобное притяжение между углеводородными радикалами, принадлежащими таким аминокислотам, как валин, лейцин или фенилаланин. Вода выталкивает эти радикалы точно так же, как вытолкнула бы обычные молекулы углеводородов, и они отлично слипаются вместе, если оказываются при сворачивании белковой молекулы близко друг к другу. А тем самым они это сворачивание и закрепляют.
В-третьих, существует электростатическое притяжение между положительно и отрицательно заряженными боковыми цепями. Если, например, глутамат (радикал которого заряжен отрицательно) окажется при сворачивании белка рядом с лизином (радикал которого заряжен положительно), между ними тут же возникнет самая настоящая ионная связь.
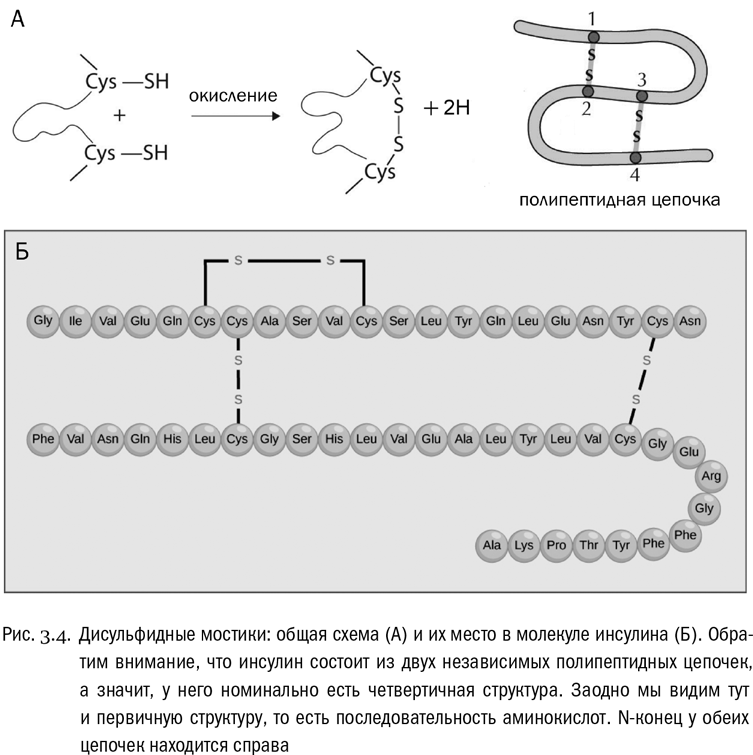
Есть и четвертый тип взаимодействий. Он зависит от единственной аминокислоты, радикалы которой могут образовать между собой аж ковалентные связи (не имеющие никакого отношения к пептидным). Эта аминокислота – цистеин. В радикале цистеина есть сульфгидрильная группа – SH, аналогичная спиртовой группе (–OH), но с атомом серы вместо атома кислорода. Целиком радикал цистеина имеет вид – CH2–SH. Так вот, уже в готовом белке может произойти реакция, при которой у двух таких радикалов будет отобран водород (его унесут специальные молекулы-переносчики), а свободные валентности атомов серы замкнутся друг на друга и образуют между остатками цистеина связь – S – S–. Это называется дисульфидным мостиком (см. рис. 3.4А). Белок вполне может быть в нескольких местах “сшит” такими мостиками (см. рис. 3.4Б). Причем реакция их образования обратима: дисульфидные мостики могут возникать и рваться, и это бывает важно для регуляции некоторых физиологических процессов.
Связи и уровни
Для удобства принято выделять четыре уровня структуры белка. Они так и называются: первичная структура, вторичная, третичная и четвертичная.
Первичная структура – это просто последовательность аминокислот, соединенных пептидными связями (см. рис. 3.4Б). Она всегда линейна, ибо белки не ветвятся. Перечислять аминокислоты в белке принято от N-конца (свободная аминогруппа) к C-концу (свободная карбоксильная группа). Множество таких перечислений, то есть записей первичной структуры белков, есть в современных электронных базах данных, доступных в сети. Можно сказать, что первичная структура белка одномерна, в то время как все остальные уровни – трехмерны. К первичной структуре относятся только пептидные связи, а к остальным уровням – любые другие взаимодействия между аминокислотами, входящими в один и тот же белок.
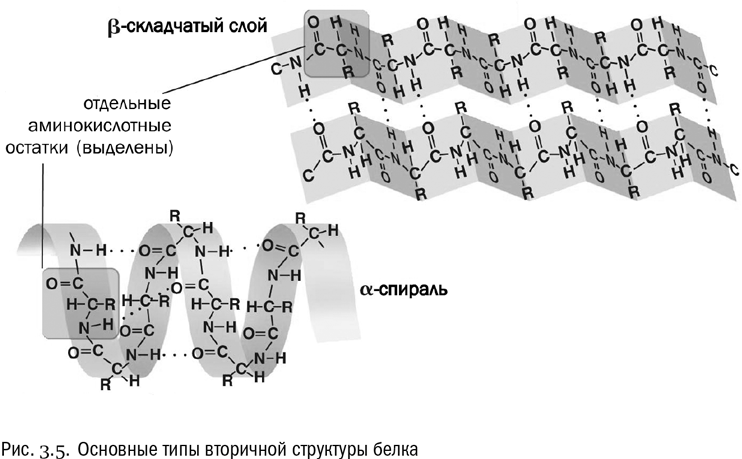
Вторичная структура – это система взаимодействий между аминокислотами в составе одной и той же полипептидной цепочки, расположенными близко (через считаные остатки друг от друга). Вторичная структура держится в основном на водородных связях (см. рис. 3.5). Причем в данном случае это связи между пептидными группами, а не боковыми цепями. А поскольку все пептидные группы одинаковы, то вторичная структура обладает высокой регулярностью, в ней часто повторяется один и тот же “узор”.
Два самых распространенных типа вторичной структуры белка – альфа-спираль и бета-слой. В альфа-спирали водородные связи постоянно образуются между аминокислотными остатками с номерами n и (n+4), то есть каждая аминокислота образует водородную связь с аминокислотой, четвертой по счету от нее. В результате получается компактная спираль, внутри которой находятся пептидные группы, а радикалы торчат в стороны. Альфа-спираль очень устойчива, в том числе и потому, что внутри нее в образовании водородных связей принимают участие все пептидные группы без исключения. В бета-слое полипептидная цепочка несколько раз перегибается, и водородные связи образуются между ее противоположно направленными отрезками.
Третичная структура белка – это система взаимодействий между сколь угодно далекими (но принадлежащими к одной и той же полипептидной цепи) остатками аминокислот (см. рис. 3.6, 3.7А). Она определяет, какую форму будет иметь молекула белка целиком. Если вторичная структура – это ближний порядок, то третичная – дальний порядок. В образовании третичной структуры участвуют водородные связи между боковыми цепями, гидрофобные взаимодействия (очень частый случай) и ионные связи между заряженными боковыми цепями. И дисульфидные мостики тоже вносят в третичную структуру свой вклад.
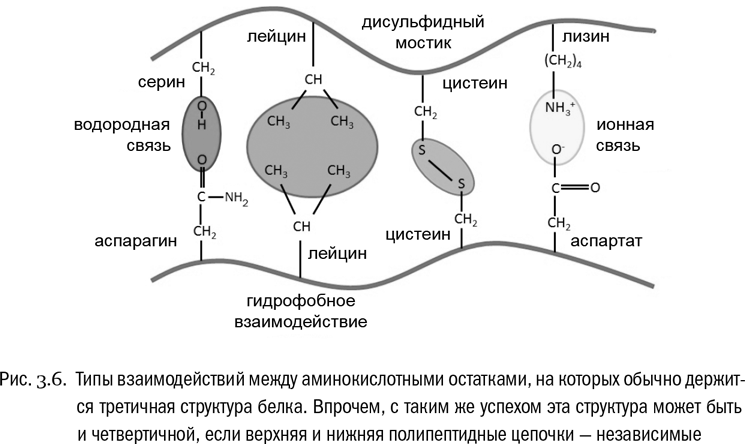
Наконец, четвертичная структура возникает в том случае, если функциональный белок собирается из нескольких отдельных полипептидных цепей (см. рис. 3.7Б). Если белок состоит из одной полипептидной цепи, значит, четвертичной структуры у него нет. Взаимодействия, создающие четвертичную структуру, те же самые, что и в третичной структуре, только не внутри одной полипептидной цепи, а между разными цепями.
Типичный белок с четвертичной структурой – гемоглобин, переносящий кислород в нашей крови. Его молекула состоит из четырех полипептидных цепочек, которые синтезируются отдельно, но свою функцию выполняют только вместе. Объединяются они в основном за счет гидрофобных взаимодействий. Всего молекула нормального гемоглобина взрослого человека включает 574 аминокислоты.
Потеря белком своей пространственной структуры без разрушения пептидных связей (то есть первичной структуры) называется денатурацией, что буквально значит “потеря природы” (см. рис. 3.7В). Самый простой способ денатурировать белок – как следует нагреть его. Именно частичная денатурация белков является основной целью любой тепловой обработки пищи. Причем иногда этот процесс до некоторой степени обратим (при кипячении молока, например). Восстановление пространственной структуры денатурированного белка называется ренатурацией. Но бывает и необратимая денатурация. Например, белок крутого яйца после полной необратимой денатурации растворенных там молекул белков становится твердым, потому что раскрученные полипептидные цепочки перепутываются между собой. Денатурация большинства белков (но не всех!) происходит при температуре 40–50 °С. Это определяет верхний температурный предел для жизни большинства земных живых существ.
Чтобы белок выполнял свою биологическую функцию, нужна, как правило, тончайшая и очень точная “настройка” его пространственной структуры. Нарушения аминокислотной последовательности тем и опасны, что они эту структуру разрушают. Например, существует генетическое нарушение, при котором в строго определенной точке одной из цепей гемоглобина глутамат заменяется на валин. Казалось бы, всего лишь одна аминокислота заменяется на другую. Но здесь это имеет неожиданно серьезные последствия. Глутамат – аминокислота, боковая цепь которой несет отрицательный заряд, валин же нейтрален и гидрофобен. Если рядом окажутся два остатки глутамата, они будут отталкиваться. А если два остатка валина, то, наоборот, слипаться. В данном случае замена глутамата на валин приводит к тому, что слипаться начинают целые молекулы гемоглобина. А это деформирует красные кровяные клетки, в которых он содержится, и вызывает тяжелую болезнь – серповидноклеточную анемию. Именно таков ее молекулярный механизм.
Конец ознакомительного фрагмента.
Текст предоставлен ООО «ЛитРес».
Прочитайте эту книгу целиком, купив полную легальную версию на ЛитРес.
Безопасно оплатить книгу можно банковской картой Visa, MasterCard, Maestro, со счета мобильного телефона, с платежного терминала, в салоне МТС или Связной, через PayPal, WebMoney, Яндекс.Деньги, QIWI Кошелек, бонусными картами или другим удобным Вам способом.
Сноски
1
Пер. А. Попова.
Комментарии
1
Broda E. The evolution of the bioenergetic processes. Pergamon Press, 1975. Перевод на русский: Брода Э. Эволюция биоэнергетических процессов. – М.: Мир, 1978.
2
Циркин Ю.Б. Мифы Угарита и Финикии. – М.: АСТ, 2003.
3
Dobzhansky T. Nothing in biology makes sense except in the light of evolution // The American Biology Teacher, 1973, V. 14, № 3, 125–129. Русский перевод этой статьи можно прочитать в сети по адресу: http://heathland.ru/111/LJ/Dobzhansky_rus.pdf
4
Фейнмановские лекции по физике. – М.: Мир, 1965.
5
Вклад легких частиц вроде фотонов и нейтрино здесь не учтен, но в современной Вселенной он в любом случае невелик (десятые доли процента).
6
Brock W. H. The life and work of William Prout // Medical history, 1965, V. 9, № 2, 101–126.
7
Caffau E. et al. An extremely primitive halo star // arXiv preprint arXiv: 1203.2612 (2012).
8
Oddo G. Die molekularstruktur der Radioaktiven atome // Zeitschrift fur anorganische und allgemeine Chemie, 1914, V. 87, № 1, 253–268. Harkins W. D. The evolution of the elements and the stability of complex atoms. I. A new periodic system which shows a relation between the abundance of the elements and the structure of the nuclei of atoms // Journal of the American Chemical Society, 1917, V. 39, № 5, 856–879.
9
Binnemans K. et al. Rare-earth economics: the balance problem // JOM, 2013, V. 65, № 7, 846–848.
10
Burbidge E. M. et al. Synthesis of the elements in stars // Reviews of Modern Physics, 1957, V. 29, № 4, 547–650.
11
Dobzhansky T. Teilhard de Chardin and the orientation of evolution // Zygon, 1968, V. 3, № 3, 242–258. Перевод этого фрагмента несколько сокращен (без потери для смысла).
12
Красилов В.А. Нерешенные проблемы теории эволюции. – Владивосток: Дальневосточный научный центр АН СССР, 1986.
13
Bracher P. J. Origin of life: Primordial soup that cooks itself // Nature Chemistry, 2015, V. 7, № 4, 273–274.
14
Vanderbilt B. Kekule’s whirling snake: Fact or fiction // Journal of Chemical Education, 1975, V. 52, № 11, 709.
15
Irwin L. N., Schulze-Makuch D. Petrolakes // Cosmic Biology, 2011, 225–251.
16
Bracher, 2015.
17
Страйер Л. Биохимия. – М.: Мир, 1984–1985 (2 тома).
18
Менделеев Д.И. Рассуждение о соединении спирта с водою, представленное в физико-математический факультет Императорского Санкт-Петербургского университета для получения степени доктора химии (1865).
19
Друг с другом они взаимодействуют за счет так называемых ван-дер-ваальсовых сил – электростатического притяжения нейтральных молекул, возникающего между мгновенными микрозарядами, которые неизбежно образуются из-за случайного характера движения электронов внутри этих молекул. Благодаря ван-дер-ваальсовым силам даже совершенно неполярные молекулы могут притягиваться друг к другу, хотя и слабо.
20
Inagaki F. et al. Microbial community in a sediment-hosted CO2 lake of the southern Okinawa Trough hydrothermal system // Proceedings of the National Academy of Sciences, 2006. V. 103, № 38, 14164–14169.
21
Budisa N., Schulze-Makuch D. Supercritical carbon dioxide and its potential as a life-sustaining solvent in a planetary environment // Life, 2014, V. 4, № 3, 331–340.
22
Whittet D. C. B. et al. Observational constraints on methanol production in interstellar and preplanetary ices // The Astrophysical Journal, 2011, V. 742, № 1, 1–10.
23
Schulze-Makuch D. Io: Is life possible between fire and ice // Journal of Cosmology, 2010, V. 5, 912–919.
24
Азимов А. Асимметрия жизни. От секрета научных прозрений до проблемы перенаселения. – М.: Центрполиграф, 2008.
25
Vickery H. B. The origin of the word protein // The Yale Journal of Biology and Medicine, 1950, V. 22, № 5, 387–393.
26
Кольцов Н.К. Физико-химические основы морфологии // Труды Третьего Всероссийского съезда зоологов, анатомов и гистологов в Ленинграде 14–20 декабря 1927 г. – Издание Главного управления научных учреждений, 1928.
27
Williams A. N., Woessner K. M. Monosodium glutamate ‘allergy’: menace or myth? // Clinical & Experimental Allergy, 2009, V. 39, № 5, 640–646.
