


Полная версия
Общая и прикладная экология
Географическая популяция охватывает группу особей, населяющих территорию с географически однородными условиями существования. Такой тип популяции занимает сравнительно большой популяционный ареал, достаточно разграничен и относительно изолирован. Различаются плодовитостью, размерами особей, рядом экологических, физиологических, поведенческих и других особенностей. Для географической популяции возможен генетический обмен, хотя он происходит реже, чем в популяциях других типов. При перекрестном скрещивании особи каждой популяции приобретают общий морфологический тип, но несколько отличающийся от соседней географической популяции, с которой регулярного контакта нет.
Популяционный ареал может уменьшаться или увеличиваться, что связано с изменением условий среды и условий существования популяции. При освобождении экологической ниши одной популяцией она неизбежно и достаточно быстро занимается другими популяциями организмов разного уровня организации.
Искусственная интродукция популяции какого-то вида организмов обычно приводит к изменению популяционного ареала аборигенных популяций, вплоть до их полного исчезновения. Это очень важно иметь в виду при создании и развитии агробиоценозов, селитебных территорий, иных искусственных объектов.
Особи в популяции могут иметь различные типы пространственного распределения, выражающие их реакции на различные факторы, такие как наличие доступной добычи (пищи), благоприятные физические условия, конкурентные реакции и др. Выделяют следующие типы пространственного распределения особей в популяции – равномерное, случайное, регулярное и пятнистое (групповое).
Равномерное распределение особей в популяции встречается в природе достаточно редко. Чаще всего оно связано с острой конкуренцией между отдельными видами, в результате которой один вид в силу ряда причин занимает практически все экологические ниши или с искусственно созданными человеком условиями (поля сельскохозяйственных растений, выпас скота и др.).
Случайное распределение встречается только в однородной среде и у видов, не имеющих склонности к агрегации. Например, распределение рыжего мукоеда Laemophloeus testaceus F. в мешке с мукой совершенно случайно, однако по мере увеличения численности его популяции распределение принимает равномерный или пятнистый характер. То же самое можно сказать о большинстве видов организмов в начальной стадии освоения ими того или иного пространства.
Распределение пятнами или группами – наиболее часто встречающийся тип пространственного распределения. Оно связано, прежде всего, с небольшими, но очень важными для организмов изменениями в окружающей среде или поведении. По Ю. Одуму (1986) групповое распределение обеспечивает популяции более высокую устойчивость по отношению к неблагоприятным условиям по сравнению с отдельной особью.
Знание типа распределения организмов имеет большое значение при оценке плотности популяции методом выборки (в случае группового размещения площадь выборки должна быть большая).
Возьмем n выборок. Среднее число особей в каждой выборке обозначим через m и получим рассеяние или дисперсию S2 по формуле
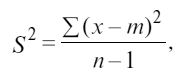
где х – фактическое число особей вида на каждой площадке. При равномерном распределении дисперсия S2 равна нулю,
так как число особей в каждой выборке постоянно и равно среднему. При случайном распределении среднее m и дисперсия S2 равны. При групповом распределении рассеяние S2 выше среднего и разница между ними тем больше, чем сильнее тенденция животных к образованию скоплений (Р. Дажо, 1975).
Численность популяции – это общее количество особей на данной территории или в данном объеме. Зависит от соотношения интенсивности размножения (плодовитости) и смертности. В период размножения происходит рост популяции. Смертность же, наоборот, приводит к сокращению ее численности.
Рождаемость – это способность популяции к увеличению числа особей за определенный период времени. Показатель характеризует частоту появления новых особей в популяции. Рождаемость определяют как число особей (яиц, семян и т. д.) – ΔN, родившихся (отложенных, продуцированных) в популяции за некоторый промежуток времени Δt. Различают рождаемость абсолютную и удельную.
А б с о л ю т н а я (о б щ а я) рождаемость – это число новых особей (ΔNn), появившихся за единицу времени (Δt).
Для того чтобы удобнее было сравнивать между собой популяции разной численности, величину ΔNn / NΔt обычно относят к общему числу особей N в начале промежутка времени Δt. Полученную величину ΔNn / NΔt называют у д е л ь н о й рождаемостью.
Поскольку в течение исследуемого промежутка Δt величина рождаемости может меняться, этот промежуток стараются сделать по возможности короче, т. е. при Δt→0 выражение ΔNn / NΔt примет вид
aNn / Ndt = b,
где d – знак дифференциала.
Полученную величину b называют также м г н о в е н н о й удельной рождаемостью. Размерность ее – «единица времени–1».
Единица времени, выбранная для оценки рождаемости в той или иной популяции, изменяется в зависимости от интенсивности размножения исследуемых организмов. Для растущей в оптимальных лабораторных условиях популяции бактерий такой единицей может быть час, для популяции планктонных водорослей – сутки, для многих насекомых – неделя или месяц, а для крупных млекопитающих – год.
Рождаемость может быть величиной положительной или равной нулю. В живых организмах заложена огромная возможность к размножению. Основная задача живого – это оставить максимальное количество потомства, что подтверждается правилом максимальной рождаемости (воспроизводства): в популяции имеется тенденция к образованию теоретически максимально возможного количества новых особей. Максимальная рождаемость является константной величиной для любой популяции.
Однако это правило может быть реализовано только в идеальных условиях при отсутствии лимитирующих экологических факторов, а размножение ограничено лишь физиологическими особенностями вида. Например, один одуванчик менее чем за 10 лет способен заселить своими потомками земной шар, если все семена прорастут, тогда как многие бактерии делятся каждые 20 мин. При таком темпе одна клетка за 36 ч может дать потомство, которое покроет сплошным слоем всю поверхность планеты. Но в реальных условиях никогда этого не происходит. Обычно действует механизм экологической, или реализуемой, рождаемости, зависящий от специфических условий среды. Поэтому максимальная рождаемость – это предел скорости увеличения числа особей в популяции.
Максимальная рождаемость как динамическая характеристика популяции важна тем, что с ней как с константной величиной можно сравнивать иные наблюдаемые величины рождаемости в тех случаях, когда известны условия среды, при которых она определяется. Наиболее корректно определение максимальной рождаемости не только при отсутствии лимитирующих экологических факторов среды, но и при оптимальных размерах популяции.
Рождаемость непосредственно связана с плодовитостью.
Плодовитость – эволюционно сложившаяся способность живых организмов компенсировать естественную смертность размножением, или скорость, с которой особь продуцирует потомков.
Характер плодовитости зависит от скорости полового созревания особей, числа генераций в течение сезона, количества в популяции самок и самцов. Если вид размножается с большой скоростью и чутко реагирует на изменения условий среды, то численность популяций его быстро и существенно изменяется.
Репродукционные возможности популяции в значительной степени определяются ее возрастной и половой структурой.
В жизни большинства организмов различают три периода: предрепродукционный (до половой зрелости), репродукционный (половая зрелость) и пострепродукционный. Относительная продолжительность их у разных видов варьируется в широких пределах, но у большинства высокоорганизованных существ обычно первый период самый длинный.
С целью описания и изучения возрастной структуры популяции используют построение пирамиды возрастов. Существует три типа пирамид:
• пирамида с широким основанием, что говорит о высоком проценте молодняка, характерная для популяции с быстрым ростом;
• со средним основанием для популяции с умеренным процентом молодняка;
• пирамида с узким основанием и численным преобладанием старых особей над молодняком, характерная для сокращающихся, угасающих популяций.
На рис. 1.3 представлены типы возрастных пирамид с их характерными особенностями.
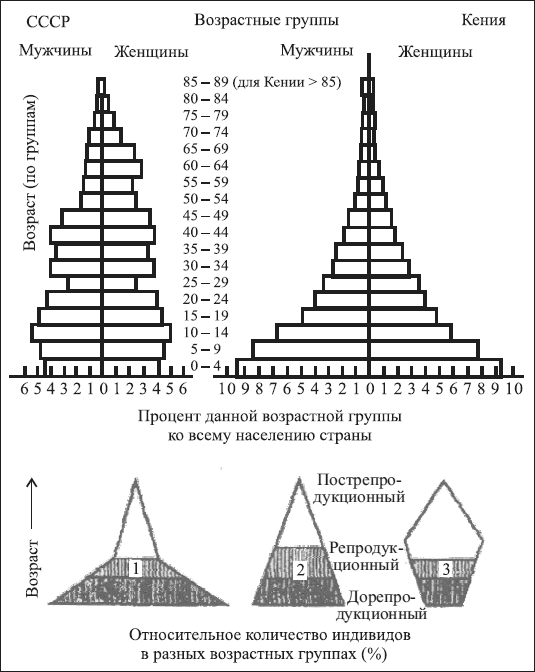
Рис. 1.3. Возрастные пирамиды и их типы (на примере населения бывшего СССР (1970) и Кении (1969): 1 – массовое размножение; 2 – стабильная популяция; 3 – сокращающаяся популяция (по Н.Ф. Реймерсу, 1990)
В 1925 г. А. Лоткой было сформулировано правило стабильности возрастной структуры популяции: любая естественная популяция стремится к стабильной возрастной структуре, четкому количественному распределению особей по возрастам. Это правило является следствием правила максимальной рождаемости (плодовитости, воспроизводства) популяции. Если это стабильное состояние из-за временного притока или оттока особей в другую популяцию по тем или иным причинам нарушается, то после восстановления прежних условий существования возрастная структура популяции будет стремиться достигнуть прежнего состояния. Наибольший успех к распространению будет иметь та популяция, которая представлена всеми возрастными группами в оптимальном их соотношении.
Правило Лотки в большей степени справедливо для высших организмов с развитой возрастной структурой популяции и не имеет свойств универсальности, однако в более широком, биосистемном, смысле оно признается универсальным.
При наличии у данного вида организмов половой дифференциации правило стабильности возрастной структуры популяции необходимо дополнять правилом стабильности соотношения полов.
Половая дифференциация (бисексуальность) играет огромную роль в поддержании генетической разнокачественности особей популяции, что, в свою очередь, обеспечивает устойчивость популяции, т. е. ее способность адекватно реагировать на изменения окружающей среды. Половая структура бисексуальной популяции определяется количеством самок и самцов в ней.
Принято выделять следующие соотношения полов в популяции: первичное соотношение полов определяется генетическими механизмами, т. е. равномерностью распределения половых хромосом; вторичное соотношение полов – это соотношение полов на момент рождения; третичное соотношение полов – это соотношение полов среди взрослых (репродуктивных) особей.
Для определения половой структуры популяции существенную роль играет половой индекс.
Половой индекс (Is) – это отношение общего числа половозрелых самок (n♀ ж) к общей численности популяции (N):
Is = n♀ … / N.
В совокупности эти два рассмотренных правила составляют правило стабильности половозрастной структуры популяции.
Таким образом, плодовитость является константой, определяемой расчетным путем, например умножением среднего числа гнезд, которые способна построить самка птицы за год, на такое же число яиц, которые она может отложить в наиболее благоприятную часть сезона года.
Смертность – величина, противоположная рождаемости, может быть определена как число особей ΔNm, погибших за время Δt. Так же как и при оценке рождаемости, смертность обычно относят к общему числу особей в популяции N, а промежуток Δt стараются брать по возможности короче.
М г н о в е н н а я удельная смертность d выражается формулой
d = dNm / Ndt.
Размерность мгновенной удельной смертности такая же, как рождаемости. Традиционно величина смертности в экологической и демографической литературе обозначается буквой d (от англ. death-rate). При этом не следует путать d (смертность) с d (знаком дифференциала в выражениях типа dN/dt).
Численность и плотность популяции зависит, наряду с рождаемостью, и от смертности.
Смертность популяции – это количество особей, погибших за определенный период. Различают абсолютную (общую) и удельную смертность.
А б с о л ю т н а я (о б щ а я) смертность – это число особей (ΔNm), погибших в единицу времени (Δt).
У д е л ь н а я смертность (d) выражается отношением абсолютной смертности к численности популяции:
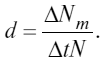
Абсолютная и удельная смертность характеризуют скорость убывания численности популяции вследствие гибели особей от хищников, болезней, старости и т. д.
Экологическая (реализованная) смертность – гибель особей в данных условиях среды. Эта величина не постоянная, она изменяется в зависимости от условий среды и состояния популяции. Полная картина смертности описывается статистическими таблицами выживаемости.
Смертность является величиной положительной или равной нулю, но последнее бывает крайне редко и только в течение короткого времени.
Различают три типа смертности:
• первый – характеризуется одинаковой смертностью во всех возрастах. Выражается экспоненциальной кривой (убывающей геометрической прогрессии). Данный тип смертности встречается редко и только у популяций, которые постоянно находятся в оптимальных условиях (человек, некоторые млекопитающие, дрозофилы и другие лабораторные животные и растения);
• второй – характеризуется повышенной гибелью особей на ранних стадиях развития и свойственен большинству растений и животных. Максимальная гибель животных происходит в личиночной фазе или в молодом возрасте, у многих растений – в стадии произрастания семян и всходов. У насекомых до взрослых особей доживает 0,3–0,5 % отложенных яиц, у многих рыб – 1–2 % количества выметанной икры;
• третий – отличается повышенной гибелью взрослых, в первую очередь старых особей. Распространен у насекомых, личинки которых обитают в почве, воде, древесине, а также в других местах с благоприятными условиями защиты, тогда как взрослые особи подвергаются постоянным воздействиям неблагоприятных факторов среды.
В связи с этим в экологии широко распространено графическое построение кривых выживания (рис. 1.4).
Располагая по оси абсцисс продолжительность жизни в процентах от общей продолжительности жизни, можно сравнивать кривые выживания организмов, продолжительность жизни которых имеет значительные различия. На основании таких кривых определяют периоды, в течение которых тот или иной вид особенно уязвим.
Если действие причин, вследствие которых изменяются рождаемость или смертность, приходится на наиболее уязвимую фазу, то их влияние на последующее развитие (рост численности, плотность, плодовитость и т. д.) будет максимальным. Это позволяет, например, регулировать численность насекомых-вредителей или грызунов в зернохранилищах.
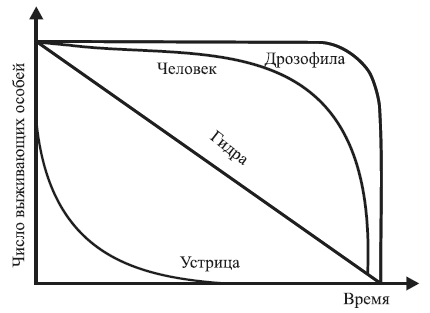
Рис. 1.4. Различные типы кривых выживания (по Р. Дажо, 1975)
Смертность подвержена более резким колебаниям и больше зависит от факторов окружающей среды, чем рождаемость, она играет главную роль в регулировании численности популяции.
Численность популяции является видоспецифическим признаком.
В любой природной системе поддерживается та численность особей в популяциях обитающих здесь животных и растений, которая в наибольшей степени отвечает интересам воспроизводства популяций.
Основное уравнение динамики численности популяции выглядит следующим образом:
r = b – d,
где r – скорость наблюдаемого изменения численности, или коэффициент прироста изолированной популяции; b – коэффициент рождаемости; d – коэффициент смертности.
В соответствии с этим уравнением рост популяции описывается экспоненциальной кривой. Это говорит о том, что оно справедливо для идеальной популяции, имеющей неограниченные пищевые ресурсы, не подвергающейся никаким внешним воздействиям среды и распространяющейся на неограниченной территории.
Понятно, что в реальных условиях эти требования не могут быть выполнены. Поэтому в 1845 г. французский математик Ферхюльст высказал гипотезу, что рост популяции человека описывает логистическая (S-образная) кривая, а в 1925 г. Р. Пирл (R. Pearl) применил эту кривую к росту любой популяции, располагающей ограниченным, но восполняемым во времени запасом пищи.
Уравнение логистической кривой выглядит следующим образом:
dN /dt = rN (K – N /K ),
где dN/dt – коэффициент роста; r – коэффициент прироста; N – численность популяции; K – максимальное число особей, способное жить в рассматриваемой среде; K – N/K – корректирующий фактор, выражающий сопротивление среды росту популяции. Отсюда
N = K (1 + ea–rt),
где а – константа, равная r/K.
Асимптота координаты K, к которой приближается кривая, соответствует пределу численности, или максимальной биотической нагруженности рассматриваемой среды (рис. 1.5).
Обычно при изучении той или иной популяции очень трудно определить ее абсолютную численность, т. е. провести количественный учет всех особей данной популяции в ареале ее обитания. Поэтому в демэкологии большей частью оперируют таким понятием, как плотность популяции.
Плотность популяции определяется количеством особей или биомассой на единицу площади либо объема (например, количество деревьев на 1 га, количество цианобактерий рода воронихия Woronichinia в 1 м3 воды).
Различают с р е д н ю ю плотность, т. е. численность или биомассу на единицу всего пространства, и у д е л ь н у ю, или э к о л о г и ч е с к у ю, плотность – численность или биомассу на единицу обитаемого пространства, доступной площади объема, которые фактически могут быть заняты популяцией.
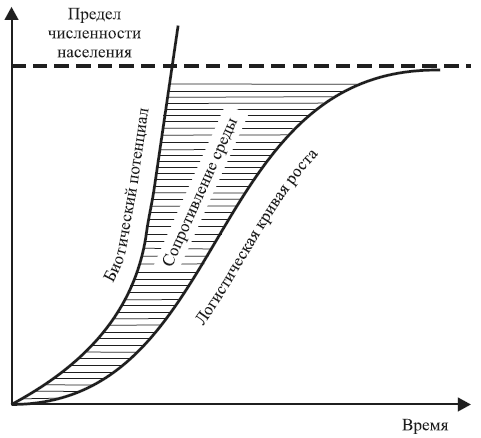
Рис. 1.5. Теоретические кривые роста популяции. Пространство, заключенное между кривой биотического потенциала Чепмана и логистической кривой Ферхюльста, соответствует сопротивлению среды (по Р. Дажо, 1974)
Плотность популяции отличается изменчивостью и зависит от ее численности. При возрастании численности не наблюдается увеличение плотности лишь в том случае, когда возможно распределение популяции за счет расширения ее популяционного ареала.
Из всего вышесказанного закономерности роста, развития и затухания популяции выглядят следующим образом:
• если рождаемость в популяции превышает смертность, то популяция, как правило, будет расти;
• с увеличением плотности скорость роста популяции постепенно снижается до нуля. При нулевом росте популяция стабильна, т. е. размеры ее не меняются. Отдельные организмы при этом могут расти и размножаться. Нулевая скорость роста означает лишь то, что скорость размножения, если оно происходит, уравновешена смертностью. Данная картина характерна для ряда одноклеточных и многоклеточных организмов, например для клеток водорослей в культуральной жидкости, для фитопланктона озер и океанов весной, для насекомых (мучные хрущаки, а также клещи, интродуцированные в новое местообитание с обильными запасами пищи, где нет хищников);
• миграция, или расселение, также как и внезапное снижение скорости размножения, способствует уменьшению численности популяции. Расселение может быть связано с определенной стадией жизненного цикла, изменениями условий среды, появлением лимитирующих факторов. Рассматривая вопрос об оптимальных размерах популяции в данной среде, необходимо учитывать кормовую продуктивность среды. Чем выше поддерживающая емкость, тем больше максимальный размер популяции, который может существовать неопределенно долгое время в данном местообитании. Дальнейшему росту популяции будут препятствовать один или несколько лимитирующих факторов. Это зависит, прежде всего, от доступности ресурсов для данного вида.
Таким образом, скорость роста популяции в естественных местообитаниях будет зависеть от климатических изменений, снабжения пищей и того, ограничено ли размножение определенным временем года и рядом других факторов.
Популяции, существующие в условиях ограниченных ресурсов, нередко хорошо подчиняются правилам логистического роста.
Численность естественной популяции может колебаться в значительных пределах.
В связи с тем что любая популяция обладает строго определенной генетической, фенотипической, половозрастной и другой структурой, она не может состоять из меньшего числа индивидов, чем необходимо для обеспечения стабильной реализации этой структуры и устойчивости популяции к факторам внешней среды. В этом и состоит принцип минимального размера популяций. Минимальная численность популяций, обеспечивающая существование вида, является специфической для разных видов.
Также относительно численности популяции, сопоставляя соотношение законов максимума биогенной энергии и давления среды, Одум (1975) формулирует правило популяционного максимума, которое звучит следующим образом: популяции эволюционируют так, что регуляция их плотности осуществляется на значительно более низкой по сравнению с верхней асимптотой емкости местообитания, достигаемой лишь в том случае, если полностью используются ресурсы энергии и пространства. При росте плотности популяции снижается обеспеченность пищей. У многих животных от потребления пищи прямо зависит плодовитость: при увеличении плотности популяции плодовитость падает, и это предотвращает дальнейший рост численности.
Правило популяционного максимума конкретизирует два обобщения. Первое из них известно как теория X.Г. Андреварты – Л.К. Бирча (1954), или теория лимитов популяционной численности: численность естественных популяций ограничена истощением пищевых ресурсов и условий размножения, недоступностью этих ресурсов и слишком коротким периодом ускорения роста популяции. Второе обобщение дополняет первое и носит название теории биоценотической регуляции численности популяции К. Фридерикса (1927): регуляция численности популяции есть результат комплекса воздействий абиотической и биотической среды в местообитании вида.
Совокупность всех факторов, способствующих увеличению численности популяции, называется биотическим потенциалом. Несмотря на то, что у разных видов составляющие биотического потенциала неодинаковы, имеется общее свойство: у всех видов он достаточно высок для стремительного увеличения численности при благоприятных условиях среды.
Рост популяции может быть столь быстрым, что приведет к популяционному взрыву. Однако следует отметить, что повышение плотности популяций сверх оптимальной оказывает на них неблагоприятное воздействие, так как при этом иссякает кормовая база, сокращается жизненное пространство, появляются эпизоотии и т. д.
Колебания численности популяции – изменение численности популяции во времени под влиянием абиотических факторов, а также в процессах иммиграции или эмиграции.
Иммиграция – активное вселение особей популяции на другую территорию.
Эмиграция – массовое выселение с занимаемой территории особей, когда плотность популяции превышает емкость среды. Различают непериодические (редко наблюдаемые) и периодические колебания численности.
Н е п е р и о д и ч е с к и е колебания численности (осцилляции) обычно носят непредвиденный характер, и причины, их вызывающие, до сих пор неясны. Р. Дажо приводит такой пример: в 1946 г. во Франции в лесу Фонтенбло жужелица Nomius pygmaeus имела среднюю численность, а жужелица Agonum quadripunctatum – очень высокую, тогда как в 1934 и 1935 гг. было выявлено всего по одной особи первого вида и несколько экземпляров второго. Причем, такая их численность держалась до 1946 г., позже ни тот ни другой вид в этом месте не встречался. Или другой пример: с 1966 г. в Тихом океане, особенно в районе Большого Барьерного рифа к северо-востоку от Австралии, началось массовое развитие морской звезды Acanthaster planci. Если до этого времени она встречалась крайне редко, то за 10 лет ее плотность увеличилась до 1 особи на 1 м2 и продолжала расти. Эта морская звезда питается полипами, составляющими живую часть коралла. Менее чем за 3 года эта морская звезда полностью уничтожила почти 40 км кораллового рифа у острова Гуам. Встал вопрос об угрозе всему Большому Барьерному рифу, что привело бы к экологической катастрофе не только в районе его нахождения, но и в других регионах планеты. Однако, через 10 лет численность морской звезды неожиданно упала до первоначальной – единичные особи на многие метры дна. Причем до настоящего времени так и не выяснено, по каким причинам происходят такие колебания численности популяций.