


Полная версия
Общая и прикладная экология
Водная среда. Гидросфера как водная среда жизни занимает около 71 % площади и приблизительно 1/800 часть объема земного шара. Основное количество воды (более 94 %) сосредоточено в морях и океанах.
В океане с входящими в него морями прежде всего различают две экологические области: толщу воды – пелагиаль и дно – бенталь. В зависимости от глубины бенталь делится на с у б л и т о р о л ь н у ю зону – область плавного понижения суши до глубины 200 м; б а т и а л ь н у ю – область крутого склона и а б и с с а л ь н у ю зону – океанического ложа со средней глубиной 3–6 км. Более глубокие области бентали, соответствующие впадинам океанического ложа (6–10 км), называют ультраабиссалью. Кромка берега, заливаемая во время приливов, называется литоралью. Часть берега выше уровня приливов, увлажняемая брызгами прибоя, получила название супралиторали.
Открытые воды Мирового океана также делятся на зоны по вертикали соответственно зонам бентали: типелигиаль, батипелагиаль, абиссопелагиаль (рис. 1.2).
Характерной чертой водной среды является ее подвижность, особенно в проточных, быстро текущих ручьях и реках.
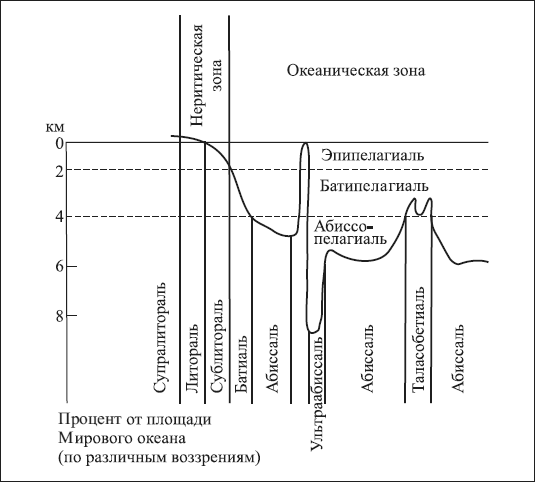
Рис. 1.2. Вертикальная экологическая зональность океана (по Н.Ф. Реймерсу, 1990)
В морях и океанах наблюдаются приливы и отливы, мощные течения, штормы. В озерах вода перемещается под действием температуры и ветра.
В водной среде обитает примерно 150 000 видов животных (около 7 % от общего их количества) и 10 000 видов растений (8 %), хотя по последним данным считается, что в водной среде могут обитать порядка 2,2 млн видов живых организмов.
Толща воды, или пелагиаль (от греч. pelagоs – море), заселена пелагическими организмами, которые обладают способностью плавать или удерживаться в определенных слоях. Эти организмы подразделяются на две группы: нектон и планктон. Третью экологическую группу – бентос – образуют обитатели дна. Организмы, располагающиеся на поверхности воды, составляют особую группу – нейстон. Организмы, часть тела которых находится над поверхностью воды, а другая – в воде, получили название плейстон.
В пресноводных водоемах различают планктон, нектон и бентос.
Водные растения в зависимости от образа жизни подразделяют на две основные экологические группы: гидрофиты – растения, погруженные в воду только нижней частью и обычно укореняющиеся в грунте, и гидатофиты – растения, которые полностью погружены в воду, иногда плавающие на поверхности или имеющие плавающие листья.
В жизни водных организмов большую роль играют вертикальное перемещение и плотность воды, температурный, световой, солевой, газовый (содержание кислорода и углекислого газа) режимы, концентрация водородных ионов (рН).
Вода является более стабильной средой, в которой ее физические параметры претерпевают сравнительно незначительные колебания, поэтому водные организмы обладают по сравнению с наземными меньшей экологической пластичностью. Пресноводные растения и животные более пластичны, чем морские, так как пресная вода как среда жизни более изменчива.
Экологическая пластичность является важным регулятором расселения организмов. Доказано, что гидробионты с высокой экологической пластичностью распространены более широко (например, элодея). А рачок артемия (Artemia solina), живущий в небольших водоемах с очень соленой водой, является типичным представителем водной фауны с узкой экологической пластичностью.
Экологическая пластичность также зависит от возраста и фазы развития организма. Например, морской брюхоногий моллюск Littorina во взрослом состоянии при отливах ежедневно длительное время обходится без воды, однако его личинки ведут исключительно планктонный образ жизни и не переносят высыхания.
У организмов водной среды выработались специфические анатомические, морфофизиологические и поведенческие адаптации к обитанию в ней.
Наземно-воздушная среда. В ходе эволюции наземно-воздушная среда была освоена позднее, чем водная, хотя в настоящее время в ней обитает значительная часть живых организмов, в том числе и человек. До настоящего времени не известно точное количество видов, обитающих в этой среде.
Особенностью наземно-воздушной среды жизни является то, что организмы в ней окружены газообразной средой, характеризующейся низкими влажностью, плотностью и давлением, высоким содержанием кислорода.
В наземно-воздушной среде действующие экологические факторы имеют ряд характерных особенностей: более высокую интенсивность света в сравнении с другими средами, значительные колебания температуры, изменение влажности в зависимости от географического положения, сезона и времени суток.
В процессе эволюции у живых организмов наземно-воздушной среды выработались характерные анатомо-морфологические, физиологические, поведенческие и другие адаптации. Например, появились органы, которые обеспечивают непосредственное усвоение атмосферного кислорода в процессе дыхания (легкие и трахеи животных, устьица растений). Получили сильное развитие скелетные образования (скелет животных, механические и опорные ткани растений), которые поддерживают тело в условиях незначительной плотности среды. Выработались приспособления для защиты от неблагоприятных факторов, такие как периодичность и ритмика жизненных циклов, сложное строение покровов, механизмы терморегуляции и др. Сформировалась тесная связь с почвой (конечности животных, корни растений), выработалась подвижность животных в поисках пищи, появились летающие животные, переносимые воздушными течениями семена, плоды и пыльца растений.
Почвенная среда. Почва (эдасфера, педосфера) – это верхняя оболочка суши, которая сформировалась в исторически обозримое время с появлением сухопутной жизни на планете. Впервые на вопрос о происхождении почвы ответил М.В. Ломоносов («О слоях земли»): «…почва произошла от согнития животных и растительных тел … долготою времени…». Великий русский ученый В.В. Докучаев (1899) впервые назвал почву самостоятельным природным телом и доказал, что почва есть «…такое же самостоятельное естественно-историческое тело, как любое растение, любое животное, любой минерал … оно есть результат, функция совокупной, взаимной деятельности климата данной местности, ее растительных и животных организмов, рельефа и возраста страны…, наконец, подпочвы, т. е. грунтовых материнских горных пород. … Все эти агенты-почвообразователи, в сущности, совершенно равнозначные величины и принимают равноправное участие в образовании нормальной почвы…».
В современной трактовке принято следующее определение почвы – это все поверхностные слои горных пород, переработанные и измененные совместным воздействием климата (свет, тепло, воздух, вода), растительных и животных организмов.
Основными структурными элементами почвы являются: минеральная основа, органическое вещество, воздух и вода.
М и н е р а л ь н а я о с н о в а (скелет), составляющая 50–60 % всей почвы, – это неорганическое вещество, образовавшееся в результате разрушения подстилающей горной (материнской, почвообразующей) породы за счет ее выветривания. Размеры скелетных частиц могут варьироваться от валунов и камней до мельчайших песчинок и илистых частиц.
Скелетный материал обычно произвольно разделяют на мелкий грунт (частицы менее 2 мм) и более крупные фрагменты. Частицы меньше 1 мкм в диаметре называют коллоидными.
Механические и химические свойства почвы в основном определяются теми веществами, которые относятся к мелкому грунту.
Физико-химические свойства почв обусловлены составом почвообразующих пород. От соотношения в почве глины и песка, размеров фрагментов зависят проницаемость и пористость почвы, обеспечивающие циркуляцию как воды, так и воздуха. В умеренном климате идеально, если почва образована равными количествами глины и песка, т. е. представляет суглинок. В этом случае почвам не грозит ни переувлажнение, ни пересыхание. И то и другое одинаково губительно как для растений, так для и животных.
О р г а н и ч е с к о е в е щ е с т в о составляет до 10 % почвы и образуется из отмершей биомассы (опад листьев, ветвей и корней, валежные стволы, отмершие травы, организмы погибших животных), переработанной в почвенный гумус микроорганизмами, некоторыми группами животных и растений.
Каждому типу почв соответствует определенный животный мир и определенная растительность. Совокупность живущих в почве организмов называют эдафоном.
Для растений имеет значение наличие достаточного количества питательных веществ в почве, влажность, кислотность (соленость), структура почвы. Эти факторы определяют видовое разнообразие и плотность распределения растительного сообщества на поверхности и верхних горизонтах почвы.
Для животных важны такие характеристики, как структура, влажность, температура почвы.
Организменная среда. Организменная среда обитания – среда, образуемая самими живыми организмами, в которых обитают другие организмы; обладает следующими особенностями:
• отсутствие света и атмосферного воздуха;
• практически постоянная температура;
• высокая влажность;
• обилие питательных веществ;
• агрессивная реакция организма – хозяина. Специфические особенности организменной среды обитания определили типы взаимоотношений организмов и обусловили особенности анатомических, морфофизиологических, поведенческих адаптаций.
Более подробно эти особенности будут рассмотрены далее в гл. 2.
1.3.2. Демэкология, или популяционная экология
Демэкология (от греч. dēmos – народ + экология), экология популяций – раздел общей экологии, изучающий динамику численности популяций, внутрипопуляционные группировки и их взаимоотношения, а также условия, при которых формируются популяции. Демэкология описывает колебания численности различных видов под воздействием экологических факторов и устанавливает их причины, рассматривает особь не изолированно, а в составе группы таких же особей, занимающих определенную территорию и относящихся к одному виду.
1.3.2.1. Популяция
Термин «популяция» был введен в экологию в 1903 г. датским ученым В. Иогансеном для обозначения «естественной смеси особей одного и того же вида, неоднородной в генетическом отношении». Он впервые применил комплекс генетических и статистических методов для изучения структуры популяции самооплодотворяющихся (самоопыляющихся) организмов. Объектом исследования стали популяции самоопылителей, которые можно было легко разложить на группы потомков отдельных самоопыляющихся растений, т. е. произвести выделение чистых линий. Анализу подверглась масса семян фасоли Phaseolus vulgaris. В настоящее время известно, что масса семян определяется полигенно и в сильной степени подвержена влиянию факторов внешней среды.
Иогансен провел взвешивание семян одного сорта фасоли и построил вариационный ряд по этому показателю. Масса варьировала в пределах от 150 до 750 мг. В дальнейшем семена массой 250–350 и 550–650 мг были высеяны отдельно. С каждого выросшего растения семена были вновь взвешены. Тяжелые (550–650 мг) и легкие (250–350 мг) семена, выбранные из сорта, представляющего популяцию, дали растения, семена которых отличались по массе: средняя масса семян растений, выросших из тяжелых семян, составила 518,7 мг, а из легких – 443,4 мг. Этим было показано, что сорт – популяция фасоли – состоит из генетически различных растений, каждое из которых может стать родоначальником чистой линии. На протяжении 6–7 поколений Иогансен отбирал тяжелые и легкие семена с каждого растения в отдельности. Ни в одной линии не произошло сдвига массы семян. Изменчивость размеров семян внутри чистой линии была ненаследственной, или модификационной.
Таким образом, Иогансен генетически неоднородные (гетерогенные) популяции противопоставлял однородным чистым линиям (или клонам), в которых невозможен отбор вследствие отсутствия выбора.
Вскоре подобные исследования были выполнены и для перекрестно-оплодотворяющихся организмов (работы Д. Джонса и Е. Иста с табаком).
Английский математик Г. Харди (1908) сформулировал понятия панмиксии (свободного скрещивания) и создал математическую модель для описания генетической структуры панмиктической популяции, т. е. популяции свободно скрещивающихся раздельнополых организмов. Немецкий врач-антропогенетик В. Вайнберг (1908) независимо от Харди создал сходную модель панмиктической популяции.
Учение о неоднородности популяций развил российский генетик С.С. Четвериков. Его работой «О некоторых аспектах эволюционного процесса с точки зрения современной генетики» (1926) было положено начало современной эволюционной и популяционной генетики. В 1928 г. А.С. Серебровский создает учение о генофонде.
В течение 1920–1950-х гг. в англоязычных странах формируется понятие идеальной популяции, и на основании этого понятия интенсивно развивается математическая генетика (С. Райт, Р. Фишер, Д. Холдейн и др.).
В нашей стране учение о популяциях развивалось в работах И.И. Шмальгаузена (популяция рассматривалась как элементарная единица эволюционного процесса), А.Н. Колмогорова (анализировались случайные процессы в популяциях) и других ученых. Однако в большинстве случаев популяция рассматривалась с общебиологической точки зрения (например, как форма существования вида – С.С. Шварц).
Лишь в 1960–1970 гг., благодаря работам Н.В. Тимофеева-Ресовского и его сотрудников формируется синтетический подход к определению популяции как эколого-генетической системы.
В настоящее время существует три основных подхода к определению понятия «популяция»: экологический, генетический и синтетический.
Экологический подход. С точки зрения экологии, популяцией является совокупность особей одного вида в пределах одного биоценоза (фитоценоза), т. е. целостная внутривидовая группировка, которой соответствует минимальная реализованная экологическая ниша. Такую группу особей иначе называют экологической или локальной популяцией, а также (для растений) ценотической популяцией или ценопопуляцией.
Р. Дажо (1975) трактует понятие «популяция» как «…совокупность особей одного вида, живущих на территории, границы которой обычно совпадают с границами биоценоза, включающего данный вид».
Ю. Одум (1971, 1975) определяет популяцию как «…группу особей одного вида (или иные группы, в которых организмы могут обмениваться генетической информацией), занимающую определенное пространство…».
Таким образом, популяция представляет собой множество особей, объединенных в пространственно-временном и экологическом отношении.
Генетический подход. С точки зрения генетики, популяция – это генетическая система, обладающая исторически сложившейся генетической структурой. Основные положения популяционной генетики возникли на основании изучения природных и модельных популяций высших раздельнополых животных (моллюсков, насекомых, позвоночных), которые воспроизводят себя с помощью нормального полового размножения – амфимиксиса, или объединения женских и мужских гамет.
В таких случаях группировка особей, способных скрещиваться между собой и производить полноценное (т. е. жизнеспособное и плодовитое) потомство, называется генетической или менделевской популяцией. В свою очередь, потомки, достигшие половозрелости, также должны скрещиваться между собой и производить полноценное потомство, т. е. популяция должна существовать длительное число поколений.
Таким образом, популяция представляет собой множество особей, объединенных достаточно высокой степенью родства. В рамках генетического подхода выделяется представление об идеальной популяции.
Идеальная популяция – это абстрактное понятие, которое широко используется в моделировании микроэволюционных процессов. При описании систем скрещивания в идеальной популяции применяют понятие панмиксии – случайного свободного скрещивания, при котором вероятность встречи гамет не зависит ни от генотипа, ни от возраста скрещивающихся особей. Если исключить половой отбор, то к панмиктической популяции применима концепция гаметного резервуара, согласно которой в популяции в период размножения формируется гаметный резервуар (генный пул), включающий банк женских гамет и банк мужских гамет. Если члены популяции равноудалены друг от друга, то встреча гамет и формирование зигот происходят случайным образом.
Реальные популяции в большей или меньшей степени отличаются от идеальной. Одним из наиболее существенных отличий является множество способов воспроизведения. По способу воспроизведения различают следующие типы популяций:
• амфимиктические – основным способом размножения является нормальное половое воспроизведение;
• амфимиктические панмиктические – при формировании брачных пар наблюдается панмиксия (свободное скрещивание);
• амфимиктические инбредные – при формировании брачных пар наблюдается близкородственное скрещивание (инбридинг, инцухт, инцест); крайним случаем близкородственного скрещивания является самооплодотворение;
• апомиктические – наблюдаются различные отклонения от нормального полового процесса (апомиксис, партеногенез, гиногенез, андрогенез);
• клональные – при отсутствии полового процесса и размножении только вегетативным путем или с помощью спор бесполого размножения (например, конидий); частным случаем клонирования является полиэмбриония – развитие нескольких зародышей из одной зиготы;
• комбинированные – например, клонально-амфимиктические при метагенезе у кишечнополостных (чередовании бесполого и полового размножения) и гетерогонии (чередовании партеногенетического и амфимиктического поколений у червей, некоторых членистоногих и низших хордовых).
Синтетический подход. В этом случае популяция рассматривается как эколого-генетическое единство признаков и свойств. Основоположники такого подхода Н.В. ТимофеевРесовский, А.В. Яблоков (1973) дают следующее определение популяции: «… это минимальная самовоспроизводящаяся группа особей одного вида, на протяжении эволюционно длительного времени населяющая определенное пространство, образующая самостоятельную генетическую систему и формирующая собственную генетическую нишу». Это определение хорошо характеризует особенности синтетического подхода.
Наиболее полным и всеобъемлющим общепринятым определением популяции является в настоящее время следующее: популяция – совокупность особей одного вида, населяющих в течение неопределенно длительного периода времени определенное пространство, внутри которой осуществляется свободное скрещивание особей (панмиксия) и которая достаточно изолирована тем или иным способом от других популяций того же вида.
В некоторых случаях при изучении популяции уместно использовать понятие формы популяционного ранга.
Формой популяционного ранга (ФПР) или группой популяционного ранга (ГПР) называют группу особей, несколько меньшую или несколько большую, чем собственно популяция. К ФПР (ГПР) меньшим, чем собственно популяция, относятся внутрипопуляционные и внепопуляционные группировки особей одного вида, которые хотя бы частично способны к самовоспроизведению. В то же время, эти группировки недостаточно изолированы от других подобных группировок, не образуют устойчивые генетические системы и не формируют собственные экологические ниши.
К ФПР большим, чем собственно популяции, относятся популяционные системы, состоящие из нескольких популяций, связанных между собой в пространственно-генетическом и (или) историческом (микроэволюционном) отношении.
Для обозначения внутрипопуляционных группировок используют различные термины: панмиктические единицы, соседства, демы и др.
Отдельно выделяют псевдопопуляции – внутривидовые группировки, неустойчивые во времени и, как правило, не оставляющие после себя потомства. Группировки популяционного ранга, внутрипопуляционные группировки и псевдопопуляции могут быть частью истинных популяций, или на их основе формируются в дальнейшем истинные популяции. Примеры таких группировок: поле пшеницы, березовая роща, колония грызунов, муравейник, население административного района (например, вороны г. Минска).
Таким образом, популяции – это надорганизменные биологические системы, которые обладают рядом свойств, не присущих отдельно взятой особи или просто группе особей. Популяция как любая сложная система характеризуется динамикой, структурой и системными (групповыми) свойствами-характеристиками.
Основные характеристики популяции. Различают статические характеристики популяции (численность, плотность, популяционный ареал) и динамические (рождаемость, смертность, относительный и абсолютный прирост численности).
Основными показателями структуры популяций является численность, распределение организмов в пространстве (популяционный ареал) и соотношение разнокачественных особей. В связи с размерами ареала популяций может значительно изменяться и численность особей в популяциях.
Популяционный ареал – территория (акватория), на которой распространена данная популяция.
П р о с т р а н с т в е н н ы й а р е а л, занимаемый популяцией, может быть неодинаковым как для разных видов, так и в пределах одного вида. Величина ареала популяции в большей мере зависит от степени подвижности особей, или радиусов индивидуальной (репродуктивной) активности. Если такой радиус невелик, то величина популяционного ареала также невелика, и наоборот. Для животных характерен еще и трофический ареал, который может не совпадать с репродуктивным. Например, белый аист Ciconia ciconia летом обитает в Европе, а зимует в Африке, т. е. обладает огромным по протяженности трофическим ареалом, так как и на месте обитания, и во время перелетов он питается по всей территории. Однако каждая пара птиц возвращается обычно в район своего старого гнезда, а популяции аистов хотя и смешиваются на местах зимовок, но во время размножения занимают достаточно небольшую территорию. Этот пример также наглядно показывает, насколько бывает трудно определить популяционный ареал и причины, его формирующие.
У растений радиус репродуктивной активности определяется расстоянием, на которое могут распространяться вегетативные части, семена или пыльца, способные дать жизнь новым особям вида.
В зависимости от размеров пространственного ареала выделяют три основных типа популяций: элементарные, экологические и географические.
Элементарная популяция (микропопуляция) – элементарная группировка особей со сходными возрастами, морфологическими, физиологическими и поведенческими показателями, приуроченные к микробиотопу (Н.В. Лебедев, 1976).
В состав их обычно входят генетически однородные особи. Количество элементарных популяций, на которые распадается вид, зависит от разнородности условий среды обитания: чем они однообразнее, тем меньше элементарных популяций, и наоборот. Между элементарными популяциями всегда имеются некоторые отличия, проявляющиеся в генетическом своеобразии, фенологических особенностях, способности к накоплению питательных веществ, интенсивности обмена, характере поведения, т. е. каждая элементарная популяция морфофизиологически и этологически (поведенчески) специфична. Различия между ними, прежде всего, определяются их генетическим своеобразием и средой обитания. Однако нередко смешение особей элементарных популяций, происходящее в природе, стирает границы между ними.
Экологическая популяция формируется как совокупность элементарных популяций. В основном это внутривидовые группировки, слабо изолированные от других экологических популяций вида, поэтому обмен генетической информацией между ними происходит сравнительно часто, но реже, чем между элементарными популяциями. Экологическая популяция имеет свои особые черты, отличающие ее в чем-то от другой соседней популяции. Белки (Sciurus vulgaris) заселяют различные типы леса, поэтому могут быть четко выделены «сосновые», «еловые», «пихтовые», «елово-пихтовые» и другие их экологические популяции.
Выявление свойств отдельных экологических популяций является важной задачей в познании свойств вида и определении его роли в том или ином популяционном ареале.