

 полная версия
полная версияMy Studio Neighbors
His labors were now concentrated on the work of interpretation, until at length his researches, covering a period of two or three years, were given to the world. In a volume bearing the following victorious title, "The Secrets of Nature in Forms and Fertilization of Flowers Discovered," he presented a vast chronicle of astonishing facts. The previous discoveries of Grew and Linnæus were right so far as they went—viz., "the pollen must reach the stigma"—but those learned authorities had missed the true secret of the process. In proof of which Sprengel showed that in a great many flowers, as I have shown at C (Fig. 3), this deposit of pollen is naturally impossible, owing to the relative position of the floral parts, and that the pollen could not reach the stigma except by artificial aid. He then announced his startling theory:
1. "Flowers are fertilized by insects."
2. Insects in approaching the nectar brush the pollen from the anthers with various hairy parts of their bodies, and in their motions convey it to the stigma.
But Sprengel's seeming victory was doomed to be turned to defeat. The true "secret" was yet unrevealed in his pages. He had given a poser to Linnæus (C), yet his own work abounded with similar strange inconsistencies, which, while being scarcely admitted by himself, or ingeniously explained, were nevertheless fatal to the full recognition of his wonderful researches. For seventy years his book lay almost unnoticed.
"Let us not underrate the value of a fact; it will one day flower in a truth." The defects in Sprengel's work were, after all, not actual defects. The error lay simply in his interpretation of his carefully noted facts. As Hermann Müller has said, "Sprengel's investigations afford an example of how even work that is rich in acute observation and happy interpretation may remain inoperative if the idea at its foundation is defective." What, then, was the flaw in Sprengel's work? Simply that he had seen but half the "secret" which he claimed to have "discovered." Starting to prove that insects fertilize the flowers, his carefully observed facts only served to demonstrate in many cases the reverse—that insects could not fertilize flowers in the manner he had declared. He was met at every hand, for instance, by floral problems such as are shown at E and F, where the pollen and the stigma in the same flower matured at different periods; and even though he recognized and admitted that the pollen must in many cases be transferred from one flower to another, he failed to divine that such was actually the common vital plan involved. It may readily be imagined that his great work precipitated an intense and prolonged controversy, and incited emulous investigation by the botanists of his time. Though a few of the more advanced of his followers, among them Andrew Knight (1799), Köhlreuter (1811), Herbert (1837), Gärtner (1844), clearly recognized the principle and foreshadowed the later theory of cross-fertilization, it was not until the inspired insight of Darwin, as voiced in his "Origin of Species," contemplated these strange facts and inconsistencies of Sprengel that their full significance and actual value were discovered and demonstrated, and his remarkable book, forgotten for seventy years, at last appreciated for its true worth. Alas for the irony of fate! Under Darwin's interpretation the very "defects" which had rendered Sprengel's work a failure now became the absolute witness of a deeper truth which Sprengel had failed to discern. One more short step and he had reached the goal. But this last step was reserved for the later seer. He took the fatal double problem of Sprengel—as shown at E and F, to express the consummation pictorially—and by the simple drawing of a line, as it were, as indicated between G and H, instantly reconciled all the previous perplexities and inconsistencies, thus demonstrating the fundamental plan involved in floral construction to be not merely "insect fertilization," the fatal postulate assumed by Sprengel, but cross-fertilization—a fact which, singularly enough, the latter's own pages proved without his suspicion.
Thus we see the four successive steps in progressive knowledge, from Grew in 1682, Linnæus, 1735, Sprengel, 1787, to Darwin, 1857-1858, and realize with astonishment that it has taken over one hundred and seventy-five years for humanity to learn this apparently simple lesson, which for untold centuries has been noised abroad on the murmuring wings of every bee in the meadow, and demonstrated in almost every flower.
This infinite field now open before him, Darwin began his investigations, and the whole world knows his triumphs. He has been followed by a host of disciples, to whom his books have come as an inspiration and ennobling impulse. Hildebrand, Delpino, Axell, Lubbock, and, latest and perhaps most conspicuous, Hermann Müller, to whom the American reader is especially referred. "The Fertilization of Flowers," by this most scholarly and indefatigable chronicler, presents the most complete compendium and bibliography of the literature on the subject that have yet appeared. Even to the unscientific reader it will prove full of revelations of this awe-inspiring interassociation and interdependence of the flower and the insect.
Many years ago the grangers of Australia determined to introduce our red clover into that country, the plant not being native there. They imported American seed, and sowed it, with the result of a crop luxuriant in foliage and bloom, but not a seed for future sowing! Why? Because the American bumblebee had not been consulted in the transaction. The clover and the bee are inseparable counterparts, and the plant refuses to become reconciled to the separation. Upon the introduction and naturalization of the American bumblebee, however, the transported clover became reconciled to its new habitat, and now flourishes in fruition as well as bloom.
Botany and entomology must henceforth go hand-in-hand. The flower must be considered as an embodied welcome to an insect affinity, and all sorts of courtesies prevail among them in the reception of their invited guests. The banquet awaits, but various singular ceremonies are enjoined between the cup and the lip, the stamens doing the hospitalities in time-honored forms of etiquette. Flora exacts no arbitrary customs. Each flower is a law unto itself. And how expressive, novel, and eccentric are these social customs! The garden salvia, for instance, slaps the burly bumblebee upon the back and marks him for her own as he is ushered in to the feast. The mountain-laurel welcomes the twilight moth with an impulsive multiple embrace. The desmodium and genesta celebrate their hospitality with a joke, as it were, letting their threshold fall beneath the feet of the caller, and startling him with an explosion and a cloud of yellow powder, suggesting the day pyrotechnics of the Chinese. The prickly-pear cactus encloses its buzzing visitor in a golden bower, from which he must emerge at the roof as dusty as a miller. The barberry, in similar vein, lays mischievous hold of the tongue of its sipping bee, and I fancy, in his early acquaintance, before he has learned its ways, gives him more of a welcome than he had bargained for. The evening primrose, with outstretched filaments, hangs a golden necklace about the welcome murmuring noctuid, while the various orchids excel in the ingenuity of their salutations. Here is one which presents a pair of tiny clubs to the sphinx-moth at its threshold, gluing them to its bulging eyes. Another attaches similar tokens to the tongues of butterflies, while the cypripedium speeds its parting guest with a sticking-plaster smeared all over its back. And so we might continue almost indefinitely. From the stand-point of frivolous human etiquette we smile, perhaps, at customs apparently so whimsical and unusual, forgetting that such a smile may partake somewhat of irreverence. For what are they all but the divinely imposed conditions of interassociation? say, rather, interdependence, between the flower and the insect, which is its ordained companion, its faithful messenger, often its sole sponsor—the meadows murmuring with an intricate and eloquent system of intercommunings beside which the most inextricable tangle of metropolitan electrical currents is not a circumstance. What a storied fabric were this murmurous tangle woven day by day, could each one of these insect messengers, like the spider, leave its visible trail behind it!
As a rule, these blossom ceremonies are of the briefest description. Occasionally, however, as in the cypripedium and in certain of the arums, or "jack-in-the-pulpit," and aristolochias, the welcome becomes somewhat aggressive, the guest being forcibly detained awhile after tea, or, as in the case of our milkweed, occasionally entrapped for life.

The Garden Sage
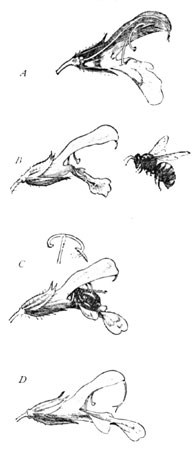
Fig. 4. Cross-fertilization of the Sage
From this companionable point of view let us now look again at the strange curved stamen of the sage. Why this peculiar formation of the long curved arm pivoted on its stalk? Considered in the abstract, it can have no possible meaning; but taken in association with the insect to which it is shaped, how perfect is its adaptation, how instantly intelligible it becomes! Every one is familiar with the sage of the country garden, its lavender flowers arranged in whorls in a long cluster at the tips of the stems. One of these flowers, a young one from the top of the cluster, is shown at A (Fig. 4), in section, the long thread-like pistil starting from the ovary, and curving upward beneath the arch of the flower, with its forked stigma barely protruding (B). There are two of the queer stamens, one on each side of the opening of the blossom, and situated as shown, their anthers concealed in the hood above, and only their lower extremity appears below, the minute growth near it being one of the rudiments of two former stamens which have become aborted. If we take a flower from the lower portion of the cluster (D), we find that the thread-like pistil has been elongated nearly a third of an inch, its forked stigma now hanging directly at the threshold of the flower. The object of this will be clearly demonstrated if we closely observe this bee upon the blossoms. He has now reached the top of the cluster among the younger blossoms. He creeps up the outstretched platform of the flower, and has barely thrust his head within its tube when down comes the pair of clappers on his back (C). Presently he backs out, bearing a generous dab of yellow pollen, which is further increased from each subsequent flower. He has now finished this cluster, and flies to the next, alighting as usual on the lowermost tier of bloom. In them the elongated stigma now hangs directly in his path, and comes in contact with the pollen on his back as the insect sips the nectar. Cross-fertilization is thus insured; and, moreover, cross-fertilization not only from a distinct flower, but from a separate cluster, or even a separate plant. For in these older stigmatic flowers the anther as it comes down upon his back is seen to be withered, having shed its pollen several days since, the supply of pollen on the bee's body being sufficient to fertilize all the stigmas in the cluster, until a new supply is obtained from the pollen-bearing blossoms above. And thus he continues his rounds.
The sage is a representative of the large botanical order known as the Mint family, the labiates, or gaping two-lipped flowers, the arched hood here answering to the upper lip, the spreading base forming the lower lip, which is usually designed as a convenient threshold for the insects while sipping the nectar deep within the tube. This mechanism of the sage is but one of many curious and various contrivances in the Mint family, all designed for the same end, the intercrossing of the flowers.
While each family of plants is apt to favor some particular general plan, the modifications in the various species seem almost without limit.
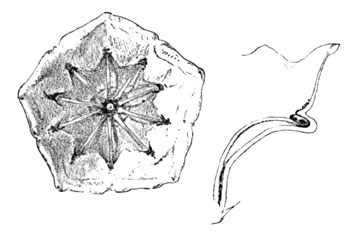
Let us now look at the Heath family. The family of the heath, cranberry, pyrola, Andromeda, and mountain-laurel—how do these blossoms welcome their insect friends? This group is particularly distinguished by the unusual exception in the form of its anthers, which open by pores at their tips, instead of the ordinary side fissures. Two or three forms of these anthers are shown in my row of stamens (Fig. 1).
Seen thus in their detached condition, how incomprehensible and grotesque do they appear! And yet, when viewed at home, in their bell-shaped corollas, their hospitable expression and greeting are seen to be quite as expressive and rational as those of the sage. Take the mountain-laurel, for instance; what a singular exhibition is this which we may observe on any twilight evening in the laurel copse, the dense clusters of pink-white bloom waited upon by soft-winged fluttering moths, and ever and anon celebrating its cordial spirit by a mimic display of pyrotechnics as the anthers hurl aloft their tiny showers of pollen!
Elastic Stamens.
Anthers Inserted in their Pockets
Every one is familiar with the curious construction of this flower, with its ten radiating stamens, each with its anther snugly tucked away in a pouch at the rim of its saucer-shaped corolla. Thus they appear in the freshly opened flower, and thus will they remain and wither if the flower is brought indoors and placed in a vase upon our mantel. Why? Because the hope of the blossom's life is not fulfilled in these artificial conditions; its natural counterpart, the insect, has failed to respond to its summons.
But the twilight cluster in the woods may tell us a pretty story.
Here a tiny moth hovers above the tempting chalice, and now settles upon it with eager tongue extended for the nectar at its centre. What an immediate and expressive welcome! No sooner has this little feathery body touched the filaments than the eager anthers are released from their pockets, and, springing inwards, clasp their little visitor, at the same time decorating him with their compliments of webby pollen (A, Fig. 5).
The nectary now drained of its sweets, the moth creeps or flutters to a second blossom, and its pollen-dusted body thus coming in contact with its stigma, cross-fertilization is accomplished. The pollen of the laurel differs from that of most of the Heath blooms, its grains being more or less adherent by a cobwebby connective which permeates the mass as indicated in my magnified representation (B, Fig. 5).
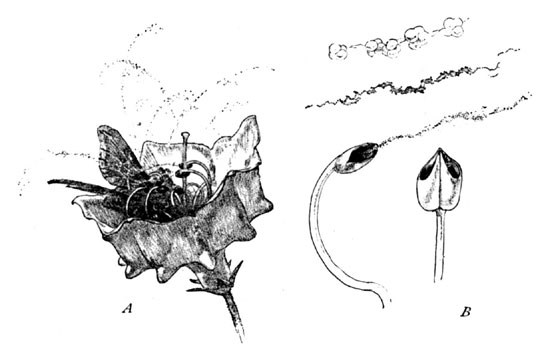
Fig. 5. Elastic Stamens of Mountain-laurel
It is probable that an accessory cross-fertilization frequently results from a mass of the pollen falling directly upon the stigma of a neighboring blossom, or even upon its own stigma, but even in the latter case, as has been absolutely demonstrated as a general law by the experiments of Darwin, the pollen from a separate flower is almost invariably prepotent, and leads to the most perfect fruition, and thus to the survival of the fittest—the cross-fertilized. And, in any event, the insect is to be credited for the release of the tiny catapults by which the pollen is discharged. But the laurel may be considered as an exceptional example of the Heath family. Let us look at a more perfect type of the order to which it belongs, the globular blossom of the Andromeda (A. ligustrina).

Andromeda Ligustrina
Only a short walk from my studio door in the country I recently observed its singular reception to the tiny black-and-white banded bee, which seems to be its especial companion, none the less constant and forgiving in spite of a hospitality which, from the human stand-point, would certainly seem rather discouraging. Fancy a morning call upon your particular friend. You knock at the door, and are immediately greeted at the threshold with a quart of sulphur thrown into your face. Yet this is precisely the experience of this patient little insect, which manifests no disposition to retaliate with the concealed weapon which on much less provocation he is quick to employ. Here he comes, eager for the fray. He alights upon one of the tiny bells scarce half the size of his body. Creeping down beneath it, he inserts his tongue into the narrowed opening. Instantly a copious shower of dust is poured down upon his face and body. But he has been used to it all his life, and by heredity he knows that this is Andromeda's peculiar whim, and is content to humor it for the sweet recompense which she bestows. The nectar drained, the insect, as dusty as a miller, visits another flower, but before he enters must of necessity first pay his toll of pollen to the drooping stigma which barely protrudes beneath the blossom's throat, and the expectant seed-pod above welcomes the good tidings with visions of fruition.
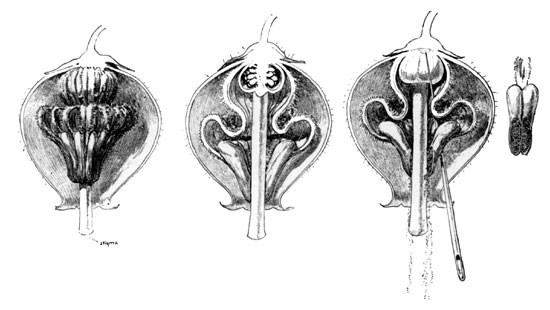
Fig. 6. Fertilisation of Andromeda

The Laurel
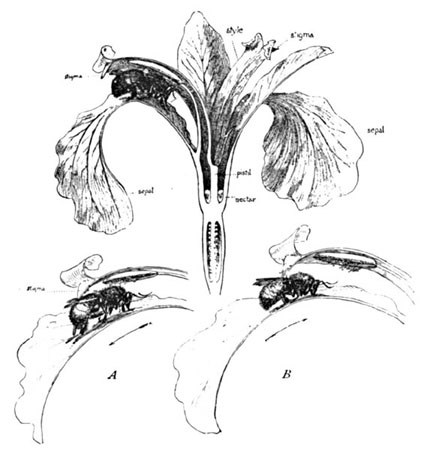
Fig. 7. Cross-fertilization of the Blue-flag

Blue-flag
And how beautiful is the minute mechanical adaptation by which this end is accomplished! This species of Andromeda is a shrub of about four feet in height, its blossoms being borne in close panicled clusters at the summit of the branches. The individual flower is hardly more than an eighth of an inch in diameter. From one of three blossoms I made the accompanying series of three sectional drawings (Fig. 6). The first shows the remarkable interior arrangement of the ten stamens surrounding the pistil. The second presents a sectional view of these stamens, showing their peculiar S-shaped filaments and ring of anthers—one of the latter being shown separate at the right, with its two pores and exposed pollen. The freshly opened blossom discloses the entire ring of anthers in perfect equilibrium, each with its two orifices closed by close contact with the style, thus retaining the pollen. It will readily be seen that an insect's tongue, as indicated by the needle, in probing between them in search for nectar, must needs dislocate one or more of the anthers, and thus release their dusty contents, while the position of the stigma below is such as to escape all contact.
In most flowers, with the exception of the orchids, the stamens and pollen are plainly visible; but who ever sees the anthers of the blue-flag? Surely none but the analytical botanist and the companion insect to whom it is so artfully adjusted and so demonstrative. This insect is likely to be either a bumblebee or a species of large fly. In apt illustration of Sprengel's theory of the "path-finder" or honey-guide, the insect does not alight at the centre of the flower, but upon one of the three large drooping sepals, whose veins, converging to the narrow trough above, indicate the path to the nectar. Closely overarching this portion is a long and narrow curved roof—one of three divisions to the style, each surmounting its veined sepals. Beneath this our visiting bee disappears, and a glance at my sectional drawing shows what happens. Concealed within, against the ridge-pole, as it were, the anther awaits his coming, and in his passage to and from the nectar below spreads its pollen over his head and back. Having backed out of this segment of the blossom (A, Fig. 7), he proceeds to the next; but the shelf-like stigma awaits him at the door, and scrapes off or rubs off a few grains of the pollen from his back (B). Thus he continues until the third segment is reached, from which he carries away a fresh load of pollen to another flower. It will be seen that only the outer side of this appendage is stigmatic, and that it is thus naturally impossible for the blue-flag to self-fertilize—only one instance of thousands in which the anther and stigma, though placed in the closest proximity, and apparently even in contact—seemingly with the design of self-fertilization—are actually more perfectly separated functionally than if in separate flowers, the insect alone consummating their affinity.
In some flowers this separation is effected, as I have shown, by their maturing at different periods; in others, as in the iris, by mere mechanical means; while in a long list of plants, as in the willow, poplar, hemp, oak, and nettle, the cross-fertilization is absolutely necessitated by the fact of the staminate and stigmatic flowers being either separated on the same stalk or on different plants, the pollen being carried by insects or the wind. We may see a pretty illustration of this in the little wild flower known as the devil's-bit (Chamælirium luteum,), whose long, white, tapering spire of feathery bloom may often be seen rising above the sedges in the swamp. Two years ago I chanced upon a little colony of four or five plants at the edge of a bog. The flowers, all of them, were mere petals and stamens (B, Fig. 8). I looked in vain for a single stigmatic plant or flower; but far across the swamp, a thousand feet distant, I at length discovered a single spire, composed entirely of pistillate flowers, as shown in A (Fig. 8), and my magnifying-glass clearly revealed the pollen upon their stigmas—doubtless a welcome message brought from the isolated affinity afar by some winged sponsor, to whom the peculiar fragrance of the flower offers a special attraction, and thus to whom the fortunes of the devil's-bit have been committed.
In some flowers this separation is effected, as I have shown, by their maturing at different periods; in others, as in the iris, by mere mechanical means; while in a long list of plants, as in the willow, poplar, hemp, oak, and nettle, the cross-fertilization is absolutely necessitated by the fact of the staminate and stigmatic flowers being either separated on the same stalk or on different plants, the pollen being carried by insects or the wind. We may see a pretty illustration of this in the little wild flower known as the devil's-bit (Chamælirium luteum,), whose long, white, tapering spire of feathery bloom may often be seen rising above the sedges in the swamp. Two years ago I chanced upon a little colony of four or five plants at the edge of a bog. The flowers, all of them, were mere petals and stamens (B, Fig. 8). I looked in vain for a single stigmatic plant or flower; but far across the swamp, a thousand feet distant, I at length discovered a single spire, composed entirely of pistillate flowers, as shown in A (Fig. 8), and my magnifying-glass clearly revealed the pollen upon their stigmas—doubtless a welcome message brought from the isolated affinity afar by some winged sponsor, to whom the peculiar fragrance of the flower offers a special attraction, and thus to whom the fortunes of the devil's-bit have been committed.
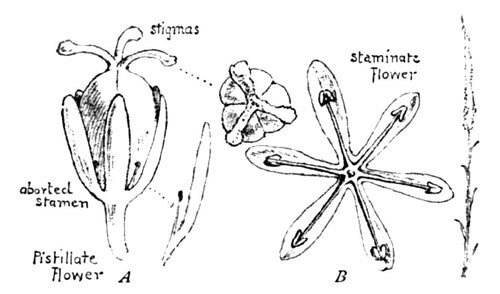
Fig. 8. Devil's-bit
The presence of fragrance and honey in a diœcious flower may be accepted in the abstract as almost conclusive of an insect affinity, as in most flowers of this class, notably the beech, pine, dock, grasses, etc., the wind is the fertilizing agent, and there is absence alike of conspicuous color, fragrance, and nectar—attributes which refer alone to insects, or possibly humming-birds in certain species.
Look where we will among the blossoms, we find the same beautiful plan of intercommunion and reciprocity everywhere demonstrated. The means appear without limit in their evolved—rather, I should say, involved—ingenuity. Pluck the first flower that you meet in your stroll to-morrow, and it will tell you a new story.

Horse-balm. Collinsonia
