


Полная версия
Нейронный мир – полное объяснение эмпирической реальности. Введение
Способность младенцев к построению силлогизмов была доказана философом Люка Бонатти [27, стр. 89], который провёл эксперимент, где младенцам демонстрировались объекты за ширмой: один из них прятали в горшок, а второй показывали младенцу, в результате чего он совершал правильный вывод об объекте, находящемся в горшке, не выражая удивления, но удивлялся, когда в горшке оказывался объект, им ранее увиденный.
Распознавание лиц начинается еще в утробе, что было доказано в одном исследовании, где младенцам подавали световые сигналы в форме лица и в форме пирамидки: реакция на лицо была более явная (Reid et al., 2014), [31]. Также младенцы уже в утробе способны различать родной и иностранный языки, что подтверждает «априорность» речи (Mehler et al., 1988), [32]. Способность младенцев к различению объектов говорит об активном развитии височной доли (Baillargeon R, 1991, 2008), [161—162].
Основные четыре столпа, на которых зиждется познание это:
1. Внимание – система нейронных сетей, которая обрабатывает и усиливает, передаёт сигналы, определяемые как релевантные: т.е. silence network (SN), переключающая внимание между экстероцепцией, интроспекцией и интероцепцией, а также dorsal attention network (DAN), участвующая преимущественно в зрительном внимании, т. е. ЭВМПНМ.
2. Активное вовлечение – подразумевает мотивацию и любопытство; обуславливается системой дофаминергической мотивации.
3. Обратная связь – коррекция АМНМ в рамках познания отношений между объектами нейронного мира, их отношения к модели субъекта.
4. Консолидация – мозг консолидирует, группирует и свёртывает, конвергирует и корректирует данные в ходе сна [27].
Вообще обучение – это и есть формирование нейронного мира (АМНМ), интернализация, происходящая посредством усиления связей между нейронами. Вычисления, производимые мозгом, не доходят до модели субъекта: они скрыты в закулисье. Только наиболее вероятный результат доходит до я-модели в качестве, например, моделирования формы тарелки, антиципации цветов, уточнения глубины, интерпретации лиц. Все эти вычисления – строгая химическая каузальность, основы которой закодированы в 750 мегабайтах ДНК кода, что, кроме всего, самоорганизуется: например, при отращивании аксонов направление движения конуса роста детерминирует филоподий, реагирующий на определённый градиент концентрации: именно так разрастается «древо» нейронного мира. Филоподий буквально вынюхивает химические метки, распластованные по коре.
Основное свойство обучения мозга – это корректировка АМНМ, заключающаяся в детализации и подстройке под параметры: при смещении, например, угла зрения с помощью специальных очков, нейросети зрительной коры начнут перестройку, в результате чего через какое-то время индивид сможет адаптироваться к соответствующему изменению. Мозг организован в виде иерархии нейронных моделей, каждая из которых ответственна за какой-либо аспект нейронного мира либо за множество аспектов. Вербальное обучение – это, в частности, регулировка «позиции вершины» при интерпретации грамматических языковых конструкций в зависимости от языковой группы (SVO – subject – verb – object), (VSO, VOS, OVS, OSV):
1. Организм реплицирует ДНК (SVO)
2. Организм ДНК реплицирует (SOV)
3. Реплицирует ДНК организм (VOS)
4. Реплицирует организм ДНК (VSO)
5. ДНК реплицирует организм (OVS)
6. ДНК организм реплицирует (OSV)
Позиция вершины детерминируется в ходе научения в раннем возрасте в зависимости от того, к какой языковой группе относится язык. Грамматика всех языков может быть описана 50 бинарными параметрами. Словарный запас в 50 000 слов укладывается в 250-килобайтный файл. В языках нет ничего особенного. Они – линейная комбинация нейронных связей. Каждый уровень обработки: от фонетического до синтаксического – не более чем иерархическая система обнаружения «закономерностей в закономерностях». В целом мозг обучается по принципу обратной связи: анализа ошибок и коррекции состояния, формирования иерархии репрезентаций, многоуровневых линейных процессов обработки данных: чем выше уровень обработки, тем большее число нейронов учится реагировать на комплексные признаки. Посредством группировки признаков происходит абстрагирование содержания. В частности, Джанет Веркер, изучившая способность младенцев обрабатывать речь, обнаружила, что младенцы способны различать звуки речи на разных языках, однако эта способность снижается в течение года, т.к. синапсы, обрабатывающие инородную речь, не активируются и, следовательно, отмирают по указанной причине. Несмотря на общее уменьшение плотности коры мозга, в течение первых 20 лет жизни речевые зоны уплотняются, что и обуславливает развитость речи у Homo s.
По ходу развёртывания нервной трубки происходит формирование основных отделов ЦНС: именно на этом этапе ДНК детерминирует все аспекты НМ, кодируя филоподии на определённый градиент концентрации: в результате дендриты разрастаются, ведомые химическими метками, и формируют НМ: уже на данном этапе он полностью создан, т.е. ничего нового в течение жизни организма сюда не прибавится. Большие полушария в случае Homo s. – вместилище НМ – вырастают из телеэнцефалического пузырька (телеэнцефалона), из чего явствует, что прозэнцефалически пузырёк эмбриона детерминирует развитие НМ [9, стр. 10]. В течение нескольких первых недель развития плода реализуется миграция нейронов, которая закладывает основу нейронной архитектуры, т. е. НМ закладывается ещё до того, как сенсорные сигналы произведут синаптические активации: например, зоны, генерирующие речь, не нуждаются в обучении у младенцев, которые только что родились – они сразу же начинают интерпретировать сенсорные данные. После образования нейроны начинают мигрировать из зоны пролиферации, формируя мозговые структуры. В больших полушариях, имеющих ламинарное строение, разрастание идёт путём деления клеток, в результате которого развивается корковая пластинка, где в будущем будет расположена большая часть нейронов: 87 млрд. клеток исходит из деления одной единственной! Одна радиальная глиальная клетка формирует 100 нейронов, простираясь по всей площади больших полушарий. Видовые различия мозга возникают из-за числа циклов клеточного деления в коре больших полушарий (БП): чем больше циклов, тем развитее БП.
На этой стадии нейроны активно взаимодействуют, закладывая основы пространства и времени: клеточная сеть, возникшая на данном этапе, является тем миром, который «оживляется» в ходе рецепторной активации стимулами. Например, паттерны разрядов клеток в зрительной системе дают сигналы, которые определяют сложную структуру латеральных коленчатых тел зрительного таламуса. Разряды, происходящие в коре зародыша, опосредуют основания НМ, корректируя структуру нейронных модулей. Однако НМ окончательно приобретает устойчивую форму лишь после рождения: с 2 до 4 месяцев плотность синаптических контактов удваивается, продолжая увеличиваться до 1 года. НМ формируется не только с помощью разрастания мозговой ткани, но и её сокращения – прунинга. Устранение излишних синаптических связей (вплоть до 100 000 в секунду) является шлифовкой НМ, подобной той, что совершает скульптор, отсекая ненужные части статуи. Пруниниг зиждется на нейронной конкуренции, которую называют нейронным дарвинизмом: самые сильные и устойчивые нейроны со множеством связей выживают, а те, что не сформировали достаточно связей, угасают. Их гибель обусловлена отсутствием поступления нейротрофических факторов (питания), например, фактора роста нейронов (NGF). Получается, что нейроны, подобно млекопитающим, борются за ограниченный объем нейротрофического фактора. Т.е. борьба происходит везде: даже на уровне мозга. В нейронах, недополучивших «питания», начинают экспрессироваться гены, запускающие смерть нейрона. Этот процесс называется апоптозом.
Рис. 12 Развитие НМ
Важным маркером хода развития мозга является степень миелинизации нейронов, обусловленная активностью шванновских клеток (ПНС) и олигодендроцитов (ЦНС). Интересно то, что некоторые зоны мозга миелинизируются позже остальных: видимо, в первую очередь подвергаются миелинизации зоны, ответственные за синтез сенсорных воздействий, а впоследствии те, что отвечают за сложные и интегративные модели. Например, развитие лобных долей, как было доказано Здравко Петанжеком, проанализировавшим плотность дендритных шипиков в ДЛПФК, у несхожих возрастных групп различается: лобные доли наиболее чувствительны к эпигенетическим факторам, а их развитие продолжается и после 20 лет. Именно лобные доли коррелируют с уровнем интеллекта: уменьшение толщины коры и рост числа связей между медиальными областями коры, латеральными областями поясной коры, латеральными областями теменной коры – основные факторы, опосредующие уровень интеллекта. «Сеть по умолчанию», образуемая дорсальной медиальной префронтальной корой (ДМПФК), задней поясной корой, предклиньем и угловой извилиной, т. е. DMN, напрямую обуславливает интеллект, т.к. отвечает за автобиографический рассказ, планирование, абстрактную модель времени (модель будущего, прошлого), теорию разума, ментализацию, социальные оценки, эпизодическую память. Таким образом, можно выделить шесть базовых векторов развития мозга: 1) морфологические изменения; 2) нейрогенез; 3) глиогенез; 4) синаптогенез; 5) миелинизация; 6) прунинг.
Подробнее рассмотрим усложнение поведения на примере 1) двигательного поведения; 2) речевого поведения; 3) способностей к решению задач. Важно иметь в виду, что соответствующее поведение не может возникнуть раньше, чем сформируется необходимый нейронный механизм, поскольку нейронный механизм и есть поведение: эта истина полностью разбивает дуализм свойств и параллелизм.
1. Ряд аксонов нейронов моторной коры миелинизируется в то же время, когда у детей 8—10 месяцев развиваются хватательные движения пальцами, а особенно – пинцетный захват, который представляет неотъемлеую часть тонкой моторики. МРТ-исследования показывают, что повышение точности движения связано с уменьшением плотности коры левого полушария, отвечающей за движение кисти у правшей (Sowell, Thomson, leonard et al., 2004), [33]. Очевидно, что разрушение данной зоны у взрослого ведёт к потере способности управления кистью.
2. К 2 годам деление и миграция клеток в речевые зоны завершается. Основной вектор «дозревания» речевых модулей состоит в синаптогенезе и миелинизации. У детей с задержкой речевого развития происходит неизбежное нарушение на одном из этапов.
3. Младенцы, по наблюдениям Пиаже, обладают априорными моделями объектов. Однако понимание физических законов, например, принципа сохранения объёма жидкости, проявляется у детей где-то в 7 лет. Поэтому несложно заключить, что априорные модели мозга в ходе его развития расширяются до более абстрактных и комплексных гипотез, что опосредует сложное поведение, детальное представление о мире. Выделяют, по Пиаже, следующие стадии развития мозга:
А) Сенсомоторную (0—2 года) – познание мира органами чувств: детализация НМ, утончение моторных навыков. Характеризуется формированием и детализацией представлений о постоянстве объектов НМ и их свойств (ЭМНМ).
Б) Дооперационную (2—7 лет) – вербализация объектов НМ, формирование я-модели, эгоцентризм, ролевые игры (ЭЯМНМ).
В) Конкретных операций (7—11 лет) – логические операции в отношении отдельных событий, антиципация аналогий, манипуляция представлениями об объектах НМ. Формируется автобиографический речевой рассказ (АЯМНМ).
Г) Формальных операций (12+) – развитие абстрактного мышления, моральных суждений, т.е. ментализации, детализация АМНМ. Существует четыре качественных скачка в развитии мозга. Каждый из них соответствует описанной стадии. Данные выводы так или иначе применимы ко всем биологическим видам, ведь НМ однороден у всех землян.
5. Я-модель (ЯМНМ)
5.1. Единство «Я» – «ТЫ»
Модель других – качественно то же самое, что и я-модель, поскольку моделируется одними и теми же нейронами по одному причинностному принципу синхронизации. Мозги всех землян способны создавать модель других, но наиболее детальна она у социальных животных и насекомых: 1) слонов; 2) дельфинов; 3) приматов; 4) муравьев; 5) пчёл; 6) ос; 7) шершней 8) и иных.
Немало страданий вызвано модулем ментализации, т.к. он предопределяет иллюзию того, что тела нечто переживают и о чём-то думают: однако ни одному телу ещё не удавалось что-либо чувствовать и переживать: всякое переживание – предсказательная модель, лишённая всякой реальности и локализованная не в теле, а в активности мозга. Иллюзия, создающая видимость того, что материальные объекты, организмы, субъекты с «психическими» состояниями, закручивает волчок страданий, ведя к деторождению, войнам, конфликтам, убийствам: все эти явления берут начало из модуля ментализации, т.е. из наивного представления о том, что другие представляют некие субъекты, которые нечто желают. Однако это мирское представление, встроенное в мозг и не подвергаемое анализу, посему безосновательно и ложно, т.е. безоснователен наивный реализм в отношении интроспекции.
Интенциональность – это абстрагированная ментализация, бредовое философское заблуждение, согласно которому «ментальное сознание» всегда на что-то направлено. Однако в рамках тоннеля НМ нечему и некуда направляться, поскольку нет субъекта, но имеется иллюзия субъективности; нет объекта, но наличествует видимость объектности. Подобные ложные и квазифилософские рассуждения исходят из деятельности мозга, конструирующей тоннель НМ, который сам себя принимает за нечто реальное и самостоятельное, закручивая колесо подкрепления и привязывая себя к самому себе. Всё в мозге, как и в организме, буквально создано для того, чтобы: 1) индуцировать НМ и поддерживать его транспарентность; 2) привязывать тоннель НМ к самому себе, чтобы обуславливать деторождение, продолжать безумство ДНК-молекулы.
В итоге вырисовывается следующая картина: земляне яростно сражаются за возможность создания нового тоннеля НМ, поддерживается который с величайшим трудом в ходе тяжкой борьбы под открытым небом, подразумевающей устранение других тоннелей НМ: их пересечение вызывает столкновение нейронных миров, что всегда заканчивается либо уничтожением одного, либо обоих. Поэтому повсеместно наблюдается пересечение тоннелей НМ и их борьба, со стороны похожая на буйство пациентов в доме умалишённых: в воде, в воздухе, на земле и под землёй, в зависимости от причудливой формы, какую приобрела материя, сформировав, таким образом, акцентуацию тоннеля НМ: на земле – ЭАМП (аудиальная), ЭВМП (визуальная), ЭОМП (обонятельная), ЭВкМП (вкусовая); под землёй – ЭКМП (кинестетическая), ЭОМП (обонятельная); в воде – ЭВМП, ЭАЭМП (аудиальная эхолокация).
5.1.2. Нарушение я-модели
Недавнее исследование «Neuronal correlates of the LSD experience revealed by multimodal…», [11], использующее нейровизуализацию для изучения эффекта ЛСД на добровольцах, продемонстрировало рассинхронизацию нейронов сети DMN, являющейся основой я-модели и ты-модели, что коррелировало с сообщениями испытуемых о растворении «Я» (self-dissolution). МЭГ обнаружила ослабление волновых альфа-ритмов.
Однако эффект ЛСД связан не только с десинхронизацией ритмов, но и со связью тех нейросетей, которые в норме работают обособленно: в итоге происходит своеобразное мультмодальное смешивание, когда одна модель НМ сливается с другой, доказывая, что НМ лишён истинного существования. Подобное «связывание» и «развязывание» нейронных модулей лежит в основе галлюцинаций. Сенсорные сигналы начинают обрабатываться не теми системами, какими должны: V1 связывается с зонами, с какими обычно не взаимодействует, чем обуславливается нарушение ЭВМПНМ (визуальной), ЭВММНМ (материи), ЭМВНМ (времени). Мозг десинхронизируется, что приводит к временному краху НМ. Это доказывает фундаментальную значимость синхронии. Поэтому заключаем, что транспарентность тоннеля НМ обусловлена правильной синхронизацией коннектома.
Агонист серотониновых рецепторов 2A (5HT) ведёт к десинхронизации ритмов: ЛСД снижает, как выяснилось ещё в 1960-х, мощность колебаний в низкочастотных диапазонах и повышает частоту альфа-ритмов (M. Finn «EEG and human psychopharmocology», 1969), [42]. Исследования воздействия псилоцибина на мозг (RL Carhart-Harris, et al. «The entropic brain»), [41] выявили снижение кровотока (CBF), снижение активности в сетях покоя (RSN) – сети DMN: отсюда выводим, что галлюциногены оказывают энтропийный эффект на мозг, повышая десинхронизацию на сетевом уровне, что подтверждает предположение, что есть нормальный уровень энтропии (НУЭ), при превышении или снижении которого НМ распадается в зависимости от степени сбоя. Эта мысль подтверждается тем («The effects of LSD-25…», 1965), [43], что электрическая стимуляция медиальных височных цепей вызывает нарушение ЭВМПНМ, т.е. визуальные галлюцинации, похожие на галлюцинации от приёма ЛСД. Из анализа 12 мозговых сетей в результате воздействия ЛСД
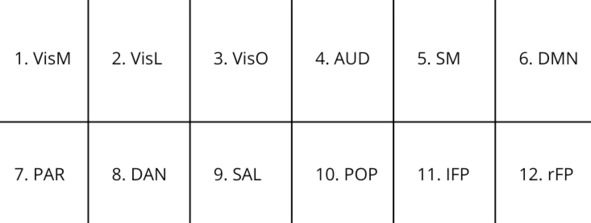
Табл. 2 Нейросети, на которые воздействует ЛСД [11]
явствует, что наиболее десинхронизация коснулась визуальных сетей (1, 2, 3) и сетей (6, 7, 8, 12).

Табл. 3 Усиление нейронный связей от ЛСД [11]
Снижение интенсивности дельта-ритмов (1—4 Гц) коррелировало с расщеплением я-модели, из чего следует, что дельта-ритм ответственен за поддержание я-модели в 1) височно-теменных областях; 2) задней части лимбической доли; 3) передней части височной доли; 4) теменно-затылочных долях.
Снижение альфа-ритмов (8—13 Гц) коррелировало с простыми галлюцинациями, т.е. сбоем в ЭВМПНМ, посему ясно, что альфа-ритм, снижающий нейронный шум, закладывает основание устойчивости ЭМНМ: 1) в затылочной зоне; 2) в височно-теменных областях. Вывод, согласно которому альфа-ритмы обеспечивают устойчивость НМ, подтверждается тем, что степень снижения альфа-частот позволяет прогнозировать интенсивность галлюцинаций.
Я-модель устойчива только в том случае, когда сети 1) DMN; 2) MPC; 3) PCC; 4) P (precuneus); 5) AG (angular gyrus – угловая извилина) работают в едином ритме колебаний, который нарушается ЛСД либо иной другой причиной, вплоть до механического устранения связи: в таком случае я-модель с неизбежностью расщепляется.
Исходя из предположения, что альфа-ритм оказывает тормозную функцию, калибруя НМ и отфильтровывая нейронный шум (O. Jensen, A. Mazaheri «Shaping functional architecture by oscillatory alpha-activity, 2010») [40], следует, что его снижение способствует зашумлению нейронных проводящих потоков, опосредуя дезорганизацию и модели мира, и я-модели в зависимости от зон, в каких возник сбой. Нарушение модели мира (ЭВМПНМ вызвано связыванием стриарной и экстрастриарной коры с зонами, которые обычно не вовлечены в создание ЭМНМ, т.е. с просачиванием АМНМ на уровень эмпирической. Альфа-канал зрительной коры, подавляясь ЛСД, разрушает ЭВМПНМ в результате того, что увеличиваются «анархические» паттерны, т.е. нейронный шум, вызывающий галлюцинации. Это подтверждает мысль о том, что чем выше энтропия на уровне нейросетей, тем интенсивнее и страдание.
5.2. Ты -модель (ТМНМ)
Ты-модель, как её называют теория разума, есть у всех млекопитающих и многих насекомых, но детальнее она у социальных. В конструировании ты-модели нет ничего особенного, подобно тому, как нет и в я-модели, что следует из однородности всех моделей НМ.
В отношении интроспективной части я-модели следует выделить её вербальный аспект – речевую интроспекцию, заключающуюся в вербальном самоотчете мозга и, соответственно, невербальную, что состоит в непосредственном моделировании иллюзии интроспекции. Поэтому первую следует назвать АВМИНМ – абстрактной вербальной моделью интроспекции НМ, а вторую – АНМИНМ (невербальной). Отсюда истекает и речевое описание ЭМНМ, которое следует назвать АРОЭМНМ – абстрактное речевое описание ЭМНМ: ЭМП, ЭММ, ЭМВ, ЭМПр. Вербализуется любая модель НМ, любой её аспект, даже непосредственно вербальная модель в виде её символизации, например, в форме логического описания высказывания.
Таким же образом ты-модель распадается на эмпирическую и абстрактную с вербальным и невербальным аспектами: эмпирическая ты-модель – это поддержание нейросетями DMN и MNS иллюзии ментализации, способности «других» переживать и думать. Она же вербализуется посредством коммуникации, т.е. любое общение представляет вокализацию АВТМНМ (абстрактной вербальной ты-модели).
Абстрактная невербальная ты-модель (АНТМНМ) – это представление о других, как о субъектах переживаний и действий, их интенциональности. Эмпирическая невербальная ты-модель (ЭНТМНМ) – это непосредственное конструирование иллюзии других в реальном времени, включающее: 1) ментализацию; 2) ЭМП; 3) ЭММ; 4) ЭМВ. Абстрактная вербальная ты-модель (АВТМНМ) – это воображаемый диалог с другими либо вокализация такового. Получается, что ты-модель распадается на: 1) ЭВТМНМ; 2) ЭНТМНМ; 3) АВТМНМ; 4) АНТМНМ.
5.3. Основание ты-модели
Ты-модель зиждется на теории разума (ТР), представляющей развивающуюся ментализацию. Теорию разума рассматривается с позиции трёх моделей: 1) модульного принципа; 2) теории теорий; 3) имитационной теории. Теория разума, покуда она теория, представляет преимущественно АМНМ с компонентами ЭМНМ, из которой она диссоциируется. Если обобщить эти наработки, то станет ясно, что моделирование «чужих» и «собственных» переживаний представляет предсказательные нейронные вычисления, работающие по байесовскому принципу, т.е. через нейронную причинность, природа которой априорна, т.е. врождённа. Выходит, что нейросети ментализации через нейронную синхронизацию моделируют как иллюзию субъектности «Я», так и субъектности «Ты» однокачественно, т.е. одна и та же нейросеть создаёт я-модель и ты-модель. Пустота предсказательного процесса познаётся следующим образом: часть пучка DMN, отвечающая за ты-модель, пытается угадать предсказательные процессы другой части пучка DMN, создающей я-модель в ином мозге. В результате наблюдается игра в предсказания: поэтому достаточно некому индивиду узнать факт из биографии другого индивида, как он сочинит историю его жизни. Именно так работают предсказательные сети ментализации. Тот, кто подобному предаётся – наивный реалист в отношении модели субъекта (ЭМСНМ и АМСНМ). Модель теории разума Барона-Коэна включает: 1) детекцию намерений; 2) распознавание направления взора; 3) разделённое внимание; 4) модуль ментализации. Детектор намерений, интегрируясь с детектором направления взора (ДНВ), конвергируется с механизмом разделённого внимания, который, в свою очередь, переходит в модуль теории разума (МТР). Все эти модули присущи всем землянам, ведь, как уже обозначалось, все нейронные миры однородны [14]. Видимо, модуль разделённого внимания зиждется на пространственных нейронах взгляда и поворота головы, поэтому в данном случае АМНМ синтезируется с ЭМНМ и как бы переходит в неё, что реализуется через синхронизацию гиппокампа и коры тета-ритмами. Выходит, что нейроны взгляда и поворота головы опосредуют эмпирическую модель разделённого внимания.
В соответствии с учением об отрицании НМ, иллюзионист подавляет ментализацию через медитацию: ты-модель, как и «Я», часть тоннеля НМ, посему лишена какого-либо истинного существования, т.к. обусловлена причинностью: физическими и химическими мозговыми процессами, а то, что обусловлено, не может существовать самостоятельно, т.е. лишено всякой самостоятельной реальности, а к тому, что её лишено, необходимо правильное отношение, как к подобному иллюзии. Иллюзионизм заключается в знании обусловленной природы всех состояний тоннеля НМ: относимся к нему как к миражу: только кажется, что он существует, но если подойти ближе и попытаться его рассмотреть, то видимость тут же исчезает. Так и остальные модели НМ: кажущаяся реальность обусловлена интенсивностью мозгового метаболизма: его снижение означает угасание НМ: при гипоксии первыми выходят из строя модель субъекта и модель тела, т.к. потребление кислорода нейронами ухудшается, из чего следует, что снижение метаболизма на уровне пучка DMN приведёт к разрушению я-модели Примерно то же происходит при десинхронизации мозговых ритмов: я и ты-модели растворяются, но по иному механизму.
Немаловажный и фундаментальный аспект ты-модели – зеркальные нейроны, осуществляющие поддержание иллюзии различия собственного тела и тел «других». Поле Бродмана 44 (Rizzolatti, 2002), [44] отвечает за моделирование движений руки и кисти в сторону определённого местоположения: отсюда следует, что зеркальные нейроны поддерживают один из аспектов кинестетической модели пространства. Система зеркальных нейронов вносит значительный вклад в поддержание иллюзии интенциональности – наличия намерений у организмов, хотя организмы качественно ничем не отличаются от любых иных материальных объектов, посему у них не может быть намерений, но возможны потребности, т.е. страдания, и поведение, определяемое синхронией нейронов и направленное на устранение страданий.
