


Полная версия
Нейронный мир – полное объяснение эмпирической реальности. Введение
3. Бета-ритм (13—35 Гц); (≤20 мкВ)
Бета-ритм, как и гамма, ответственен за конструирование ЭМНМ: способен подавлять АМТНМ. Например, бета-ритм моделирует ЭВМПНМ и конкурирует с альфа-римтом, который её подавляет. Дело в том, что нативная ЭЭГ бодрствования представляет смесь периодических и непериодических паттернов, поэтому все ритмы действуют в строгой взаимосвязи. Ретикулярная активирующая система (РАС) ингибирует бета-ритм при пробуждении, что соответствует активации ЭМНМ, поэтому бета-ритм ответственен, по крайней, мере за усиление ЭМНМ. Холинергические нейроны (ХН) базальных отделов переднего мозга, синтезирующие ацетилхолин, запускают формирование бета-ритма бодрствования при пробуждении ото сна, т.е. индуцируют ЭМНМ: ХН проецируются в кору, где запускают каскад реакций, приводящий к ингибированию медленных ритмов и ускорению быстрых. В свою очередь, ЭМТ опосредуется серотонинергической системой, которая формирует «ритмы движения». Бета-ритм следует считать связывающей мозговой осцилляцией, синтезирующей сенсорные модальности (Howe et al. 2011; Takahashi et al. 2015), [62], а также ритмом, организующим модель поведения (Leventhal et al. 2012), [63]: т.е. данный ритм более выражен при включении организма в решение конкретной задачи, нежели при спонтанном поведении (Mallet et al. 2008), [64]. Из этого следует, что бета способен оказывать стабилизирующее действие на ЭМНМ, снижая нейронный шум (Engel and Fries 2010), [65].
1) Аудиальная бета-синхронизация:
Сихронизация мозгом ритма музыкального с двигательными паттернами осуществляется бета ритмом за счёт связывания моторных и аудиальных зон (ЭМТ – ЭАМНМ), (Merchant and Yarrow 2016), [66]. Интенсивность бета-ритма во время прослушивания ритмического произведения снижается после тона и увеличивается перед началом следующего, в результате чего реализуется аудиальное кодирование (Fujioka et al. 2009), [67]. Бета-активность наблюдается в слуховой коре в том случае, когда ритм регулярный, и исчезает при нерегулярном (Fujioka et al. 2012), [68]. При этом наблюдается циклическая обратная связь между моторной и аудиальной корой во время прослушивания ритмической музыки посредством бета-ритма (Fujioka et al. 2015), [69]. Таким образом, аудиомоторная система координируется бета-ритмом, предсказывающим музыкальную последовательность. Из этого следует, что бета-ритм предопределяет ЭАМНМ.
2) Конструирование ЭМВНМ (модель времени):
Бета-ритм вносит немалый вклад в производство ЭМВНМ. Модель времени, опосредованная отчасти бета-ритмом, задействуется мозгом для синхронизации двигательных актов (Teki 2014), [70], а также в ходе аудиального кодирования [66]. Закономерно предположить, что временное кодирование (ЭМВНМ) – расширение изначально необходимого для выживания двигательного кодирования: т.е. детальная модель времени явилась в качестве следствия организации поведения мозгом и нужна для детализации этого поведения, поскольку ритм, конструирующий ЭМВНМ, позволяет реализовывать качественно более спрогнозированное поведение, а также учитывать его следствия в рамках временной проспективы (АМВНМ).
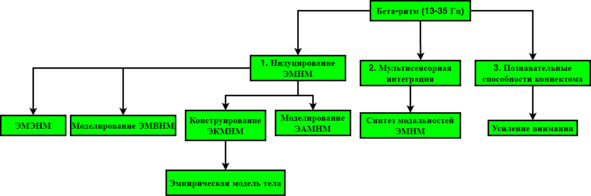
Рис. 7 Бета-ритм
3) Кинестетическая бета-синхронизация:
Всё предуготовлено к тому, чтобы признать фундаментальную роль бета-ритма в связывании двигательных последовательностей (Jin and Costa 2010), [71]; (Fujii and Graybiel 2003), [72]: из этого исходит более основополагающий механизм временного моделирования. Высвобождение двигательных паттернов из M1 осуществляется бета-ритмом (Hussain SJ, Vollmer MK, 2022), [106]. В сравнении с мю-ритмом, локализованным в S1, бета-осцилляции наблюдаются в первичной моторной коре M1, демонстрируют точную соматотопическую организацию и распространяются рострокаудально в коре (Stolk A, Brinkman L), [109]. Корково-мышечная когерентность наиболее интенсивна в бета-диапазоне, что указывает на связующую функцию бета-ритма, а также на наличие связи M1 со спинальными мотонейронами (Romei V, Bauer M), [110]. Переход от низкой фоновой активности M1 к высокой соответствует высвобождению двигательной программы, что соответствует превышению двигательного порога (Hanes DP, Schall JD, 1996), [111]. Этот процесс обусловлен активацией нейронов M1 таламокортикальными проекциями, вызывающими реализацию двигательных паттернов (Takahashi N), [112]. Интересно, что сенсомоторные ритмы (ЭМТНМ) ослабляются при воображении движения и его подготовке (АМТНМ) (Pfurtscheller G, Brunner C, 2006) [113]. Это подтверждает истинность предположения о конкуренции ЭМНМ и АМНМ.
4) Эффект резиновой руки
У индивидов с иллюзией самоприкосновения в ходе эксперимента резиновой руки наблюдалась более мощная бета-синхронизация, нежели у тех, у кого иллюзии не было (Faivre N, Dönz J, 2017), [78]. Одновременно с этим снижалась интенсивность альфа-диапазона в сенсомоторной и премоторной коре (Oakes et al., 2004), [79]. Также подавление альфа-волн обнаружилось 1) в случае иллюзии владения телом (Evans et al., 2013), [80]; (Kanayama et al., 2007, 2009), [81—82]; 2) в случае иллюзии владения пальцами.
5) Конструирование ЭМНМ
Активное конструирование ЭМНМ коррелирует со снижением альфа-активности, сильных межполушарных связей (Spadone et al., 2015), [73], (Betti et al., 2013), [74]. В то же время повышается активность бета и гамма-осцилляций, что указывает на их значимость в создании ЭМНМ: это закономерно, поскольку только быстрые ритмы обладают достаточным расширением для поддержания ЭМНМ (Cole et al., 2014), [75]; (Merchant H, Bartolo R., 2018), [76]; (Kim et al., 2017), [77]; (Hasson U, 2004), [100]. Сходство активности бета-диапазона в состоянии естественного зрения и просмотра фильма указывает на однокачественное моделирование ЭМНМ в обоих случаях, в то время как динамика альфа-ритма не изменялась в этих двух ситуациях.
4. Гамма-ритм (30—600 Гц); (≤15 мкВ)
Под гамма ритмом в данном случае имеются в виду все быстрые ритмы. Гамма-ритм детерминирует люцидный сон, посему ясно, что именно он отвечает за активное моделирование ЭМНМ (Scarpelli S, Bartolacci C, 2019), [83]; (Voss U, 2014), [103]: очевидно, что если гамма-ритм индуцирует ЭМНМ в люцидном сне, то и в бодрствовании реализует ту же задачу, поскольку нет иного механизма. То есть высокое разрешение быстрых ритмов – основа детальности ЭМНМ, поскольку медленные ритмы недостаточны для подобной задачи. Основное доказательство зависимости ЭМНМ от гамма-ритма заключается в его особой роли в мультисенсорном связывании (Bhattacharya et al., 2002), [84]; (Mishra et al., 2007), [85]; (Balz et al., 2016), [86]: это согласуется с данными о том, что гамма-ритм отражает изменения восприятия, т. е. ЭМНМ. Гамма-диапазон усиливает визуально-моторную связь, т.е. конвергирует ЭВМПНМ с ЭКМНМ (Womelsdorf et al., 2006), [87], поддерживает интеграцию распределённых в коре нейронов во время зрительного процесса (Rodriguez et al., 1999), [88], а также мультисенсорную обработку (Roelfsema et al., 1997), [89]. Гамма в этом плане осуществляет связывание соматосенсорных нейронов со зрительными (дорсальный поток).
За соматосенсорное владение иллюзорными конечностями также отчасти ответственен гамма-ритм (Engel et al., 2001), [90], а именно за прогностический аспект ложных конечностей. Продемонстрирована способность гамма-ритмов в отношении зрительного связывания, т.е. синтеза ЭВМПНМ (Tallon-Baudry, Bertrand, 1999), [91]: через 100 мс после предъявления зрительного стимула наблюдается гамма-всплеск в стриарной коре. Исходя из этого предполагается, что гамма-ритм синхронизирует зрительные карты, т. е. ЭВМПНМ. Активное исполнительное внимание усиливается гамма-волнами, что подтверждает их вклад в познавательные способности мозга [13, стр. 408].
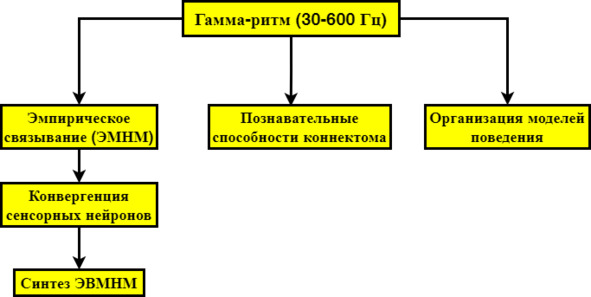
Рис. 8 Гамма-ритм
Синтез ЭМНМ
Определённо можно утверждать, что гамма-ритм реализует связывание сенсорных модальностей в объекты ЭММНМ: разные группы нейронов кодируют различные аспекты ЭВМПНМ, и мозг обладает механизмом их объединения. Синтез лиц (ЭМЛНМ) задействует гамма-связывание (Lachaux et al., 2005), [92]. Гамма-ритм сглаживает контрастные грани и ганглиозные точки света, создавая иллюзию целостности и чёткости. ТМС в затылочной зоне способна вызывать фосфены на уровне нижних гамма-ритмов (20 Гц), что указывает на его роль в т.н. доминантных комплексах нейронов, образующих ГРП (Kanai et al., 2008), [93], т.е. осознавание объектов ЭМНМ. Явление фосфенов обусловлено обратной связью от зоны MT к зоне V1. ЭМРНМ (модель речи) также детерминируется гамма-ритмом: прослушивание, восприятие и проговаривание слов представляет гамма-всплески в различных областях мозга, посему ясно, что речь образуется гамма-ритмами (Crone et al., 2006), [94].
Следует подчеркнуть системообразующую роль гамма-ритма в поддержании ЭВМНМ (Ni J, Wunderle T, 2016), [104]; (Womelsdorf T, Schoffelen JM, 2007), [105].
5. Ритмы – общие замечания
А) Учитывая то, что мысли о роли ритмов в синтезе НМ, высказанные ранее, способны ошибаться, важно отметить следующее: даже если предположение о вкладе основных ритмов в синтез НМ, описанное здесь, ошибочно, то это только подтверждает многоаспектность и многоуровневую сложность электромагнитной активности мозга. Тем не менее очевидно, что только этот механизм способен синтезировать НМ: в связи с этим истинна гипотеза, что быстрые ритмы активно синтезируют ЭМНМ, тогда как медленные представляют основание этого синтеза, группируя быстрые волны.
Б) Эксперимент Мишеля Жуве (Michel Jouvet, 1972), [95] с кошками, которым повредили (неэтично) нейроны голубого пятна, продемонстрировал следующее поведение: спящие кошки бегали и пытались поймать некий объект (мышь), очевидно, делая это в ЭМНМ, что активировалась в REM-сне, поскольку голубое пятно не индуцировало атонию мышц: то, что кошка делала в осознанном сне, не подавлялось блокировкой мышц, поэтому активность соматосенсорной и моторной коры приводила мышцы в действие, как если бы кошка охотилась на мышь с открытыми глазами в бодрствовании. Это подтверждает тот факт, что в осознанном сне активируется НМ тот же, что в бодрствовании, поскольку в ином случае подобного поведения наблюдать не удалось бы. Осознанный сон индуцируется гамма- и бета-ритмами (Voss, U., Holzmann, R., 2014), [96]; [8, стр. 245]. Из этого с очевидностью выводим, что в осознанном сне, который правильнее назвать REM-активацией ЭМНМ (R-АЭМНМ), возможна индукция той же модели мира, что тождественна бодрствованию, т.к. между двумя этими состояниями нет качественной разницы. Во время REM-сна, в рамках которого возможна индукция ОС, ацетилхолин побуждает таламус и кору вести себя так, как в бодрствовании. Также наблюдается снижение отправки сигналов системами верхнего ствола мозга – голубым пятном и ядрами шва [8, стр. 258]. Пример катаплексии доказывает, что в рамках атонического приступа нейроны голубого пятна ингибировались, а нейроны крупноклеточного ядра – индуцировались, вызывая состояние REM-сна: последние связаны с ЭМНМ. Орексиновая система также участвует в поддержании ЭМНМ, что подтверждается случаем нарколепсии, т.е. сбой орексиновой системы запускает каскад реакций, подавляющий ЭМНМ (Mieda, 2017), [97]. Это связано с тем, что гипокретиновая система – одна из модуляторных систем, локализованная в латеральном гипоталамусе и поддерживающая состояние бодрствования, т. е. ЭМНМ. Переход из не-REM-сна в бодрствование соответствует активации промежуточных нейронов таламуса, переходящих из дельта-частот в односпайковый режим, посредством 1) ацетилхолиновой стимуляции; 2) норадреналиновой стимуляции; 3) гистаминовой стимуляции: в переходе нейронов таламуса в активное состояние проявляется суммарный эффект медиаторов [8, стр. 253].
В) Отключение ретикулярной активирующей системы (РАС) равнозначно подавлению ЭМНМ, а её активация, задействующая бета-ритм, запускает ЭМНМ. Полное отключение ЭМНМ – медленноволновой сон – начинается только тогда, когда РАС снижает интенсивность, а ритмы, как следствие, десинхронизируются.
Г) Теменная кора осуществляет гамма- и бета-интеграцию сенсорных стимулов, конвергируя ЭМП, ЭММ, ЭМВ. В свою очередь, височная доля в большей степени опосредует АМНМ, в частности, ААМНМ, которая проявляется в семантизации нейронных объектов и их отношений, а также ЭВММНМ, проводя детализацию объектов НМ (Beauchamp, 2005), [98]. Островок, спрятанный височной, лобной и теменной долями, – место синтеза интерорецепции, т.е. ощущения внутренних органов тела ЭМТ.
Д) Таламус и полушария
Таламус, являясь связующим центром мозга, безусловно, ответственен за ритмическое регулирование полушарий: правильность сенсорной связности, модуляция сенсорных потоков – основание для ЭМНМ в её раздроблении. Исходя из того, что ЭМНМ в основном складывается в теменных и височных долях, следует, что области таламических проекций этих зон вносят немалый вклад в поддержание транспарентности НМ и его ритмического регулирования, придавая каждой области необходимый ритм и конвергируя различные зоны в синхронизирующем ритме: в частности, эту мысль подтверждает альфа-таламическая гипотеза Бергера. Зрительная часть таламуса индуцирует альфа-ритм, подавляя ЭВМНМ – ЛКТ. Нейроны релейных ядер таламуса посредством пейсмейкерной активности вызывают ритмическую генерацию ПД, формируя т.н. «кортикальных двойников», т.е. через таламокортико-таламическую петлю таламус достигает синхронизации с корой, реализуя модель возвратного альфа-торможения. Ретикулярное ядро регулирует кортикальный вход, осуществляя распределения альфа-ритмов в коре посредством возвратного торможения (Бреже М., 1979), [61].
Передняя часть теменной доли связана с вентральным латеральным ядром (ВЛЯ), средняя – с вентральным постлатеральным ядром (ВПЯ), а задняя – с латеральным задним ядром (ЛЗЯ), (Standring, 2005), [99]: из этого следует, что моторная кора и соматосенсорная кора связаны с ВЛЯ и ВПЯ соответственно. ЛКТ – ЭВМНМ, а МКТ – ЭАМНМ: первое принимает проекции из ГКС, а второе – из волосковых клеток базилярной мембраны. Передне-задне-боковое ядро конвергирует соматосенсорную и моторную информацию – ЭМТ.
Е) В меньшей степени подвергаются апоптозу нейроны, участвующие в сетях синхронизации (Nicolas Heck, Antje Golbs, 2008), [101]: это достигается за счёт расположения клетки, когда синаптические связи наиболее устойчивы, что повышает жизнеспособность последней. Нейроны организуются таким образом, чтобы наиболее эффективно складывать НМ (Downes JH, Hammond MW, 2012), [102].
2.4. Эмпирическая и абстрактная модели НМ (ЭМНМ и АМНМ)
Нейронный мир создаётся двумя базовыми моделями: эмпирической моделью НМ (ЭМНМ) и абстрактной моделью НМ (АМНМ). Первая представляет активное гамма- и бета-моделирование, формирующее созерцаемую реальность; вторая – менее интенсивная активация тех же зон мозга, опосредующих ЭМНМ, реализующая абстрагирование эмпирического содержания. ЭМНМ можно условно назвать «восприятием», однако мозг не контактирует с объективной реальностью, т.е. это не корректно, а АМНМ – воображением, что допустимо. На данный момент известно, что ЭМНМ ингибируется ретикулярной активирующей системой (РАС): когда организм пробуждается ото сна, РАС активирует ритм бодрствования в коре через гипоталамус, т.е. активирует НМ, а когда засыпает – ритм подавляется [8, стр. 252]. Фундаментальная роль РАС в поддержании НМ подтверждается тем, что при повреждении ствола мозга наступает непоправимая кома [5, стр. 544].
В соответствии с этим каждая модель НМ распадается на эмпирическую и абстрактную, закладывая фундаментальную дихотомию, которая условна, поскольку в действительности весь нейронный мир однороден. Однако на условном уровне мы признаем подобную дихотомию, т.к. нарушение АМНМ, т.е. включение её в пространство ЭМНМ (ГРП – глобальное рабочее пространство), представляет галлюцинации. Подобный сбой обусловлен, как и всякий сбой нейронного мира, десинхронизацией электромагнитных ритмов [1, стр. 159].
2.5. Абстрактная модель нейронного мира (АМНМ); связь с ЭМНМ
1. Природа АМНМ
АМНМ – это воображение, т.е. активность тех же зон, что опосредуют ЭМНМ. Воображение движений – это менее интенсивная активация двигательных и соматосенсорных областей, чем при движении; воображение звуков – менее интенсивная активность вторичных аудиальных областей, т. е. A2; воображение вкуса – аналогично: однако иногда воображение, т. е. АМНМ, способно перейти в ЭМПНМ, как в случае с представлением чего-то кислого, после которого возникает ощущение оскомины [23].
Во всяком мозге, создающем НМ, есть как эмпирическая, так и абстрактная модели НМ: исключительно их верное взаимодействие, обусловленное электромагнитной синхронизацией, опосредует транспарентность (прозрачность) НМ. Если ЭМНМ задаётся высокочастотными ритмами, то АМНМ – низкочастотными: например, АМПНМ, (абстрактная модель пространства НМ) в гиппокампе опосредуется тета-ритмом [13, стр. 415]. Эмпирическая модель относится к абстрактной, как закат к его пейзажу, эскизу. Без абстрактной модели бесполезна эмпирическая, и наоборот: без первой невозможно ни целенаправленное поведение, ни какое-либо познание, ни ориентация в нейронном пространстве, т.к. эмпирическая модель пространства имеет значение только тогда, когда детализируется нейронами решётки гиппокампа – АМПНМ. Поэтому очевидно, что АМНМ есть у насекомых, иначе их поведение было бы дезорганизовано.
АМНМ располагает как врождённым, так и сформированным компонентами: демонстрация силуэтного изображения кошки котятам заставляет шестинедельных котят вздыбить шерсть, оскалить зубы, реализовать модель разрешающего поведения, хотя до этого момента они никогда не видели такой позы, какую представляет силуэт [12]. Это доказывает, что в мозге котят и других животных АМНМ уже сформирована в момент рождения. В ней уже пребывают базовые элементы модели материи (ММ). Объекты, социальный контекст: позы кошек, последовательности звуков (мяуканье) и иные объекты. Получается, что мозг котят сопоставляет стимулы среды с набором врождённых критериев, т.е. происходит переход информации ЭМНМ – АМНМ.
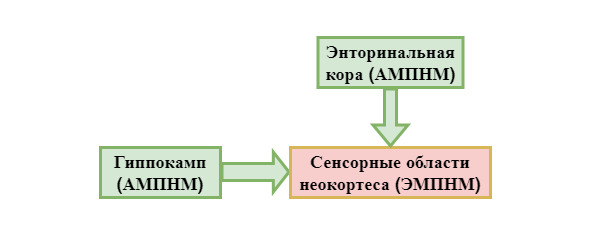
Рис. 9 Связь АМПНМ с ЭМПНМ через тета-ритм
В мозге кошек наличествует нейронный механизм, запускающий реакцию преследования и убийства, который зиждется на сопоставлении эмпирической модели объекта – жертвы, т. е. ЭММНМ, с его абстрактной формой в АМНМ. Мышь, созданная мозгом кошки в ответ на сенсорные стимулы, активированная ими, идентифицируется через вентральный зрительный поток нейронами височной доли, формирующими АМНМ как жертва, и запускается реакция погони и убийства, основанная на опиатной системе подкрепления [13, стр. 260]. Выходит, что АМНМ – это своеобразный набор критериев, с которым мозг сравнивает эмпирические объекты, им созданные. Каждый биологический вид, как правило, реагирует запуском половой программы на особь собственного вида, что также доказывает врождённость полового поведения: это сообщает и то, что конструкция половых органов уже заложена в АМНМ организмов, т.к. их вид запускает реакцию возбуждения, а значит, эмпирическая модель половых органов была идентифицирована как подходящая через височную кору, а затем запустилась нейросеть, ответственная за двигательную программу совокупления.
Врождённость АКМНМ (кинестетической) доказывается и слепыми детьми, которые демонстрируют те же выражения лица, что наблюдаются у взрослых особей: эти дети не могли видеть выражения лиц и имитировать их. Врождённость как всего НМ, так и АМНМ, объясняется закреплением адаптивных форм поведения в ходе естественного отбора: поведение – это нейронная активность, а она детерминирована кодом ДНК, посему априорность НМ и его моделей очевидна.
Ясно, почему вид Homo s. так жесток и агрессивен: сила, агрессивность и ловкость необходимы для победы над противником и, следовательно, удачной передачи ДНК. Дуэли – пример закрепления социального статуса: победившие передавали гены. Поэтому, как это ни печально, убийства, войны, насилие и иные аффективные формы поведения никогда не искоренятся, поскольку адаптивны, посему передаются по наследству: они встроены в АМНМ, например, разделение на «своих» и «чужих».
Если крысу в клетке бьют током, то она тут же нападает на ту, что невинно находилась рядом: мозг крысы связывает причину боли с ближайшим объектом НМ, формирует довольно примитивную интерпретацию произошедшего. Это напоминает ситуацию, когда машина марки «X» врезалась в толпу и уничтожила 8 человек, а затем разъярённые организмы разбивали все машины данной марки: примитивный механизм ассоциации объектов НМ управляет поведением.
Если лысая обезьяна страдает, то её мозг связывает это страдание с лысой обезьяной из другого сообщества, группы, клана, страны: так начинаются религиозные и мировые войны. Их причина – неудовлетворённость организмов текущим положением дел, т.е. страдание, а попытка удовлетворения всегда провальна, т.к. зиждется на порабощении и насилии, отбивании территорий и эксплуатации.
Поистине: могли бы организмы воевать, если бы были счастливы и довольны собственным существованием? Малое страдание от тягот существования перерастает в масштабное – кровавую баталию: так не смерть ли – истинная цель всех землян? И не муки ли – путь?
2. Иллюзия сущностей
Иллюзия мира идей, вещей в себе, абсолютов и прочего – продукты абстрагирования ЭМНМ. Шопенгауэр, объясняя природу творчества, писал, что оно зиждется на усмотрении общего в частном и познании платоновских идей. Также он подчёркивал тождество кантовской вещи в себе и платоновской идеи. Он оказался прав, но только в ином плане: не в их действительном существовании, а в их общей природе.
В действительности нет ни нематериальных идей Платона [169, стр. 271], ни вещей в себе Канта, лежащих за пределом эмпирической реальности [167], но есть абстрактная модель каждого эмпирического объекта НМ, диссоциированная корковыми сетями. Именно эти абстрактные модели создают иллюзию возвышения над частным, сжатия эмпирических объектов, выхода за пределы мира-явления. Абстракции не выходят за пределы пространства и времени, становясь вечными идеями и вещами в себе, как считал франкфуртский затворник, а представляют их части, являясь сетями функционирующего мозга. Причём имееются в виду не части нейронного пространства и времени как модели НМ, а объективное пространство и конкретные нейросети.
Поэтому всё, что пребывает в АМНМ, – нейронные выжимки эмпирических объектов ЭМНМ и их состояний, не более. Основание любых абстракций следует усматривать именно в АМНМ, а не в неких внепространственных и вневременных пластах вечности.
3. Нейронная основа АМНМ
АМНМ следует разделить на имплицитную, т.е. на входящую в глобальное рабочее пространство (ГРП), к которой относятся нейроны пространства, и эксплицитную, являющуюся воображением, различными видами памяти и опосредуемую височной, лобной и энторинальной корой; программы двигательного поведения, формирующиеся базальными ганглиями, но не имеющие проекций в кору больших полушарий и не доходящие до я-модели.
Поток сенсорных данных сначала идёт в парагиппокампальную область, затем в энторинальную кору и, в конце концов, в гиппокамп. Именно этот процесс производит диссоциацию сенсорного содержания ЭМНМ, формируя АМНМ, распадающуюся на многие модели: весь этот процесс не входит в ГРП, поэтому его качество невозможно отследить интроспективно, подобно тому, как можно отследить качество и точность движений. Посему, если этот процесс нарушается, то выпадение АМНМ замечается не сразу, а в контексте: например, различные формы ретроградной амнезии, проявляются которые нередко только в случае неудачной попытки нечто вспомнить. Гиппокамп осуществляет обратные связи в медиальные области височных долей, где происходит формирование абстрактных моделей объектов, затем в сенсорные области неокортекса,
устанавливая значимую связь АМПНМ (пространства) – ЭМПНМ.
Сенсорные данные, проходящие через лобную, теменную, височную, затылочную и поясную кору, попадают: 1) в периренальную кору, что образует сеть АММНМ (материи), что занимается идентификацией объектов ЭМНМ, информация о которых поступает непосредственно от зон, моделирующих объекты, т.е. по вентральному потоку из зрительной коры; 2) в парагиппокампальную кору, объединяющую ЭМПНМ и позволяющую распознавать сцены [13, стр. 264]; её повреждение ведёт к неспособности распознания пейзажей, «целостных» пространственных пластов, что указывает на её роль в формировании АМПНМ (пространства); 3) в энторинальную кору, содержащую а) нейроны решётки; б) нейроны границы; в) нейроны направления головы, которые проецируются на нейроны места гиппокампа (Zhang S.-J., Ye J., Couey J. J.), [20], опосредуя сообщение между ЭМПНМ неокортекса и АМПНМ в гиппокампе, где происходит более точная абстрактная локализация положения модели тела в модели пространства посредствам нейронов места: т.е. проекции из энторинальной коры на нейроны места осуществляют детализацию и поддержание транспарентности МПНМ (модели пространства).
