


Полная версия
How to Grow a Human
Now, this is certainly one way to talk about the evolution and origin of sex, but it invokes an uncomfortable amount of teleology. Sex doesn’t really exist in order to create genetic diversity. Nothing happens in evolution in order to produce a particular end result. It makes intuitive sense for us to speak like this, but the fact is that sex evolved because those early organisms that became able to fuse their cells and chromosomes somehow produced more robust populations than those that lacked this ability. Sexual reproduction might be a more or less inevitable consequence of evolution by natural selection, once it gives rise to a particular kind of complex organism, much as snowflakes are an inevitable consquence of the laws of physics and chemistry playing out in a particular environment. Evolutionary biologists say that sex is a successful evolutionary strategy, although this again imputes a sort of foresight to evolution that it doesn’t possess.
While these arguments for the value of sex surely have a lot going for them, they can’t be the whole answer. Sex is not essential for all higher vertebrates. Parthenogenesis occurs in many different types of animal, including insects such as mites, bees and wasps, and some fish, reptiles and amphibians. In a few such cases, reproduction can happen either with sex or without. Sometimes that’s by design – for example, parthenogenesis is thought to be an option for mayflies as a defence against a lack of males. (The same useful trait arises in the women of the male-free society in Charlotte Perkins Gilman’s utopian feminist novel Herland (1915).) In other cases it occurs by accident, unfertilized eggs just happening rarely to develop into embryos. Komodo dragons are among the larger creatures that can reproduce this way.
The reasons and mechanisms for parthenogenesis are varied and sometimes rather complicated. But one thing you can say for sure is that, in organisms for which it can take place, evolution has not seen fit to rule it out. To put it another way, there is nothing obviously necessary about sex, and assessing the benefits of sexual reproduction over other methods of propagating is a subtle and perhaps context-dependent business.4 As far as evolution is concerned, it is just a matter of whatever works.
* * *
There’s a complication with sex. Each of our body cells has two sets of chromosomes, and therefore dual copies of each gene, one inherited from each parent. But if one of these cells in a female simply fused with one from a male, the resulting cell would have four sets of chromosomes. That’s too many, and the cell couldn’t function properly. So organisms that reproduce sexually have evolved special kinds of cells that possess only one copy of each chromosome. These are the gametes, and they are found only in the gonads: the ovaries and testes.
Gametes are made from a specialized type of cell called a germ cell. The germ cells have doubled chromosomes (they are said to be diploid) just like somatic cells, but in a special kind of cell division called meiosis they divide into gametes in which these chromosomes have been carefully segregated into two. Cells with just a single set of chromosomes are said to be haploid.
Normal cell division (mitosis) involves replication of the chromosomes accompanied by their separation so that each daughter cell receives the full complement. Meiosis is even more complicated, because the existing chromosomes have to be divided precisely in two and shipped off to their respective destinations.
Actually it’s worse than that. Meiosis happens in two stages, and the overall result is that a single, diploid germ cell replicates its chromosomes once and divides twice to end up with four haploid gametes. As in mitosis, the process by which the chromosomes are divided uses a spindle-like structure made from fibrous protein. The chromosomes become attached to the fibres and are drawn towards opposing poles of the spindle located in the two lobes of the dividing cell.
Crucially, the chromosomes undergo some shuffling in this process. The pre-meiosis germ cell, recall, has one of each of the 23 types of chromosome from the mother, and a second copy of each from the father. Which of the poles of the spindle each chromosome is drawn towards is random, and so the diploid cells made by division of the germ cell have a random combination of maternal and paternal genes.5 The haploid gametes that eventually emerge from the process then have a thoroughly scrambled single set of chromosomes: with 23 pairs of chromosomes in all, there are 223, or about 8 million, possibilities. These are combined with a similar range of options in the other gamete when egg and sperm unite, so you can see that having sex is a good way to produce genetic diversity.
Formation of the so-called primordial germ cells happens early in the development of a human embryo, around two weeks after fertilization. This is even before the gonads have started to form, which is to say, before the embryo has yet “woken up” to which sex it is. It’s as if the embryo is putting these cells aside while deferring the matter of whether they will be eggs or sperm. The gonads themselves will guide this process, sending out chemical signals that tell the primordial germ cells which sort of gamete to become. They’re ready to do that around week six of gestation, by which time the germ cells have migrated across the developing embryo to their destination. For yes, that development involves not merely cell division but also cell movement, a physical sorting in space to arrange the parts in the proper disposition.
Germ cells were first postulated by the German zoologist August Weismann in his 1892 book The Germ-Plasm: A Theory of Heredity. As that title suggests, this was a hypothesis as much about evolution as about embryology. The “plasm” here reflects the widespread notion, before Boveri and Sutton’s chromosomal theory of inheritance, that heredity was somehow transmitted via the “protoplasm” substance inside cells. As we saw earlier, Charles Darwin speculated that the particles responsible for inheritance, which he called gemmules, were collected from the body’s cells and transmitted via sperm and egg. Weismann was a staunch advocate of Darwinism, but he was convinced that there was a fundamental distinction between the somatic cells that made up the body’s tissues and the special cells called germ cells that gave rise to gametes. Any changes to the “plasm” of somatic cells could therefore play no part in heredity. To demonstrate that changes to the body of an organism are not inherited, Weismann cut off the tails of hundreds of mice and followed their offspring for five generations, each time removing the tails. Not once were any offspring born without tails.6 Any notion that “acquired characteristics” could be inherited, as in the pre-Darwinian theory of evolution proposed in the early nineteenth century by Jean-Baptiste Lamarck, could no longer be sustained.
In Weismann’s view, then, somatic cells are irrelevant to evolution. They are destined to die with the organism. But germ cells beget more germ cells – there is an unbroken line of germ cells (the germ line) down through the generations. It’s often said that the germ cells are thus immortal, although that’s an odd formulation – by that definition, we are all immortal simply by virtue of being able (if indeed we are) to produce offspring.
* * *
In the story of how to make a human “the natural way”, the fertilized egg is often portrayed as the end – at least, until the happy day that the baby emerges. All our traditional stories of people-making rely on that quantum leap from fertilization to birth. The dire moral warnings about pregnancy that loomed over adolescence (and in some cultures still do) make this the equation: bring together sperm and egg and you’ll get a baby! It’s a warning (sometimes needed, for sure) to experimenting teenagers, but becomes more like a promise in the narrative of IVF: to make that longed-for baby, all you need to do is unite the gametes. And if it doesn’t turn out that way, something has gone wrong. There is a single and inevitable road from fertilized egg to infant, and anything else is an aberration.
This is misleading. To put it starkly, most acts of non-protected penetrative sexual intercourse do not produce a baby – and when I say most, I mean 99.9 per cent. Even most fertilized eggs do not become babies – about 2 to 3 in 10 confirmed pregnancies abort spontaneously in miscarriage, but even those figures mask the 75 per cent or so of fertilized eggs that never get to the point of registering as a pregnancy at all, either because they don’t develop into a multi-celled embryo or because the embryo fails to implant in the uterus. That’s a puzzling thing about humans: we are unusually poor, within the animal kingdom, at reproducing. You have to wonder whether all the attention we give to sex is because we are so spectacularly bad at getting results from it.
Even to say “bad” is perhaps to collude with the moral imperative of the fertilized-egg-to-infant story; let’s just say that we are an anomaly, for reasons imperfectly understood. This calls into question the idea that sex really is “for” reproduction, as some religious moralists insist. If we were inclined to see procreation as a divine gift and imperative, one would at least need to grant that God expects us to have a heck of a lot of rehearsal.
The baby grows, of course, from a fetus: even children’s books tell us that. But in the common view the fetus is simply a baby – a person – that has not yet fully developed. Its proportions might be a little odd, its limbs blunter, but it is recognizably human. The classic images made in the mid-1960s by Swedish photographer Lennart Nilsson and presented in the book A Child Is Born (1965), have defined the view of our in utero existence ever since. They show the fetus floating freely in space, often lacking even an umbilical cord, like the iconic image from Stanley Kubrick’s 2001: A Space Odyssey three years later. Perhaps this “child” even sucks its thumb. But these images were actually made by artful arrangement of aborted fetuses – they were not in fact living organisms at all, much less in utero. They were curated to tell a reassuring story. (At least, so it might seem until you realize that it’s a story in which the mother has been edited out.)
By the time a fetus looks even vaguely human (which is what, loosely speaking, distinguishes it from an embryo), most of the important stuff has happened. Most of the dangerous hurdles have been cleared. And most importantly, the developing organism is already anthropomorphic, relieving us from any need to grapple with the strangeness of an entity evidently made of cells, which we might want to call human but would struggle to justify that intuition.
Yet it is the early embryo that reveals the true versatility, the genius, of our cells – and the unfamiliarity of the moment when those cells are not merely what we are made of, but what we are.
It might surprise you to discover – it surprised me – that when a woman first has a fertilized egg (a zygote) inside her body she is not technically pregnant. This is not some perverse biomedical fine print; it simply makes no sense to see things otherwise. A pregnancy test would show nothing, nor will it for the first four days or so after fertilization. The zygote divides by mitosis into two, then four, then eight cells and so on, and at this point these cells can form all the tissues needed in the embryo: they are called stem cells and are said to be totipotent.
In other words, every one of these cells could potentially become a separate embryo. In the early days of embryology, that was by no means clear. The German zoologist Wilhelm Roux thought, for example, that cells are headed towards different fates from the first division of the zygote. In 1888, he reported experiments on frog embryos at the two and four-cell stage, in which he destroyed one of the cells by lancing it with a hot needle. A single remaining cell from a two-cell embryo would then, he said, grow into a half-embryo, suggesting that it had even at that stage become assigned as the progenitor of that part of the body plan alone.
But Roux’s method was flawed, because he could not detach the remains of the ruptured cell from the intact one. This debris interfered with the subsequent growth of the embryo. In the 1920s and ’30s, German embryologist Hans Spemann performed a cleaner act of surgery on salamander embryos. By using a noose made from a single hair taken from a baby, he pinched early embryos in two and found that each of the resulting parts is able to grow into a complete embryo.7 In effect, Spemann made identical twins by artificial means. Because he produced two genetically identical embryos from a single initial one, you could also call this a process of cloning.8 Spemann and his co-workers used amphibian cells, because they are so large that the delicate manipulation could be done by hand – albeit an impressively steady one.
The ball of totipotent stem cells that is the human embryo floats freely in the fallopian tube (also called the oviduct), borne slowly towards the uterus. By day five, the embryo has become a ball of around 70 to 100 cells and has rearranged itself into a structure known as the blastocyst. By the time it arrives at the uterus, it has shed the protein coat called the zona pellucida that formed the protective shell of the original egg – it has “hatched” and is ready to implant.
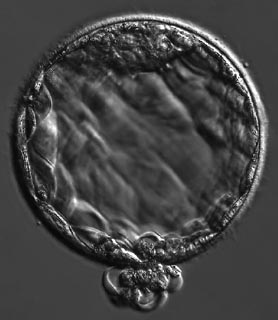
The human embryo at around five days, called a blastocyst.
That ball of cells is not exactly the nucleus of a person. Most of the cells of the blastocyst became the mere housing and life support. Some of them form an outer layer enclosing a fluid-filled void: these are trophoblast cells, comprising the tissue called trophectoderm which will become the placenta. Others congregate into a clump on the inside, called the inner cell mass, which separates into the epiblast from which the fetus will grow, and the hypoblast that will eventually become the yolk sac. The epiplast consists of embryonic stem cells, capable of forming all the tissues of the body (but not the placenta): a capacity called pluripotency. Identical twins grow from two separate inner cell masses in a single blastocyst, whereas non-identical twins grow from two separate blastocysts, formed from distinct eggs fertilized by different sperms. Within a few days of implanting, the epiblast is covered in a layer of specialized cells called the primitive endoderm, derived from the hypoblast.
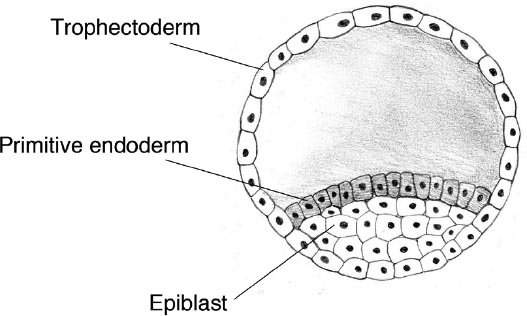
The human embryo at around day 10–11.
The fate of the embryo wholly depends on a successful implantation in the lining of the uterus. If this does not happen – which is the case around 50 per cent of the time – the embryo will be expelled in the menstrual cycle. Failure to implant is one of the common reasons why an IVF cycle does not work. No wonder, then, the division of labour in the blastocyst makes it seem that its priority is to those cells surrounding the epiblast, which won’t be a part of the fetus at all. For without implantation, it’s game over.
Implantation is a delicate and complex process involving a dialogue of hormones and proteins between the embryo and the cells of the uterine lining. In some ways it is more delicate and complex than fertilization itself. The placenta, for example, is made not just from the trophoblast layer of the blastocyst but also from tissues from the mother, called the decidua. The two types of cell, with different genetic makeup, have to work together to create a single, vital organ. Emotive and anthropomorphic metaphors suggest themselves, presenting implantation as an intimate collaboration between the tissues of mother and her “child”. But one might equally choose to speak of the blastocyst “invading” the uterine tissue: one “organism” colonizing another for its survival.9 Both are stories; neither is a neutral description of events (which story ever is?).
* * *
The best is about to come. Calling the part of the embryo fated to become the baby an “inner cell mass” is no euphemism: it really does seem to be a shapeless conglomerate. If we want to insist that baby-making is a miracle, what seems truly miraculous is not just that the inner cell mass makes a body but that, most often, it makes exactly the same type of body, with five fingers on each hand, with all facial features in the right place and fully functional, and with its battery of correctly positioned organs. It’s no surprise that development of the embryo occasionally goes awry; it is astonishing that it does so rather rarely.
When embryos start off as single cells, they have no plan to consult. Cells are programmed to grow and divide, but it isn’t meaningful to think of a human being as somehow fully inherent in a fertilized egg, any more than one can regard the complex convolutions of a towering termite mound as being programmed into each termite. The growth of an organism is a successive elaboration of interactions within and between cells: a kind of collaborative computation whose logic is obscure and convoluted, and the outcome of which is incompletely specified and subject to chance disturbances and digressions.
In this way, the job evolution has devised for those formative cells is an architectural one: a challenge of coordination in time and space. They have to move into position, to acquire the right fate at the right time, and to know when it is time to stop growing or to die.
Developmental biologists talk of this as “self-organization”. It could make the process sound quasi-magical, calling as it does upon the image of the cell as an autonomous being with aims and purposes. But many of the rules are now broadly understood.
Two key factors are at work. First, as the cells divide and multiply, they take on increasingly specialized roles, a process called differentiation. Thus, totipotent cells in a two or four-cell embryo become trophoblasts or the pluripotent stem cells of the epiblast. The latter go through further stages of differentiation that ultimately produce the specialized cell types found in muscle, skin, blood and so forth. We will see shortly how that happens.
Second, particular spatial arrangements may arise from cells actively moving through or across the growing organism or organ, or becoming sorted into clumps of different cell types by preferential stickiness, often between cells that are alike.
That cells have adhesive qualities joining them into aggregates was suggested in the 1890s by Wilhelm Roux. He was also able to disrupt frog embryos by vigorous shaking, which separated them into single cells. He found that those cells would join back together, which he attributed to some kind of attractive force.
Such “disaggregation” experiments were taken further in the early 1900s by marine biologist Henry V. Wilson, who found that sponges kept for a long time in an aquarium became “loose” and could be teased apart into individual cells. He achieved the same thing in fresh sponges by the simple measure of squeezing them through a piece of silk, which acted as a sieve that separated the cells. Again, those cells would reassemble if brought into contact to regenerate a living sponge. It was like a recapitulation of the evolution of primitive multi-celled organisms from colonies of single-celled ones (see the First Interlude, here). When Wilson did the experiment with different species of sponge, he found that cells from the same species would stick together selectively. Ernest Everett Just discerned in the 1930s that the reason for this selectivity had something to do with the cell membranes. The truth is that cells adhere via protein molecules protruding at their membrane surface (especially those belonging to the class called cadherins), which will bind to one another discerningly.
This notion of “tissue affinity” was developed around the same time by the German-American embryologist Johannes Holtfreter. In 1955, he and Philip Townes studied how the cells of amphibian tissues that had been disaggregated by exposing them to alkalis could reassemble from solution. Holtfreter largely outlined the concept of cell sorting that allows tissues of several cell types to adopt particular structures and arrangements.
The process of body formation (morphogenesis) is orchestrated by genes, and no wonder then that genes have been attributed such determinative power. Some researchers have made more apt comparisons to a musical score: genes tightly constrain but do not fully prescribe the performance. This is still a limited metaphor, because you can look at the score and figure out (if you’re a musician) pretty much how things will go. Not so with genes. Sometimes it is better simply to tell the story as it is, as simply as you can, rather than trying to pretend it is some other story.
Morphogenesis literally means shape-formation, but equally it is a question of cell specialization: the embryonic stem cells gradually lose their versatility as they divide, becoming geared instead to do the task of specific tissue types. Heart muscle cells must execute synchronized beating, pancreatic cells must secrete insulin, the nerve cells of the eye’s retina must respond to light, and so on. This happens not by cells gaining new properties, but rather by narrowing the possibilities inherently available to them by shutting down genes that aren’t needed. That’s what differentiation is all about.
The cells must know how and where to switch genes on and off as differentiation proceeds. How do they know? The cues come from the other cells and tissues around them.
Some of these signals are delivered as chemical messages, which, diffusing through the mass of cells, serve to define a kind of spatial grid that lets cells know where they are in the overall embryo and thus what their fate should be.
Imagine that a cell, or group of cells, at one place in the embryonic mass switches on a gene that produces some protein. And suppose that this protein can diffuse out of the cell, like water leaking out of a paper bag, and into other cells. Then the concentration of the protein throughout the embryo varies gradually from place to place, being greater nearest the cells that produce it and slowly diminishing with distance. If you could measure the protein concentration, you’d have some notion of where you are in the embryo relative to the source cells. You’d be able to sense your position. Think of it in the same way as finding your way to the kitchen of a large house by following the smell: the stronger it is, the closer you are.
These “position-marker” proteins are called morphogens, and cells are able to “sense” their concentrations. Morphogen concentration gradients allow regions of the embryo to become distinct from one another.
To see how this can work, let’s forget the human body for a moment and look at the development of a simpler embryo: that of the fruit fly. This humble creature became the paradigmatic representative of “complex life” in the early twentieth century, when its robustness and ease of breeding made it the ideal subject to study the mechanisms of genetic inheritance – an art of which Thomas Hunt Morgan was the master. There are, of course, substantial differences between humans and fruit flies, extending to their genetic and developmental fine print. In particular, fruit-fly embryos, unlike those of mammals, are not initially clusters of separate cells at all. Once fertilized, the ovoid fly egg starts to replicate chromosome-carrying cell nuclei, but just accumulates these around the edges of the egg. The nuclei only acquire their own cell membrane once the embryo has amassed 6,000 or so. This lack of cell membranes in the early embryo makes it particularly easy for morphogens to diffuse through it.
One simple way that gradients of diffusing molecular morphogens can mark boundaries is to think in terms of concentration contours. A contour denotes a threshold: a point where the concentration exceeds a certain value.


