


Полная версия
Боль в спине. За кулисами лечения и реабилитации. Часть III. Механизмы «неспецифической» боли
Механизмы развития «простой» неспецифической боли я разобью на отдельные составляющие/механизмы для удобства восприятия. Очень часто они встречаются одновременно, усиливая эффект и затрудняя выявление ведущего.
Основным виновником неспецифической боли являются мышцы, связки и капсулы суставов – ткани, имеющие наибольшую иннервацию свободными нервными «болевыми» окончаниями. Принято считать, что чем обильней иннервирована ткань, тем больший вклад в развитие болевых феноменов она вносит. Для позвоночника в этом плане первое место занимают капсулы суставов, второе – задняя продольная связка и диск, и третье – менискоиды – прокладки внутри сустава по типу менисков в коленном суставе. Обильная иннервация плотной оформленной соединительной ткани (фиброзной) не случайна. Данный тип ткани относится к брадитрофным, т.е. таким, для которых характерен низкий уровень питания. Соответственно, при повреждении – очень низкая репаративная способность. Поврежденный сухожильный и связочный аппарат может не восстанавливаться годами. Обильная иннервация [выполняющая сигнальную функцию], таким образом, защищает соединительную [хрящевую] ткань от возможного регулярного повреждения. Незначительная травма приводит к выраженной боли и обездвиживанию (шинированию) поврежденного участка. Другого способа защиты соединительной ткани у организма нет.
С мышцами в этом плане проще: они хоть и восстанавливаются с некоторым включением фиброзной ткани, но выраженной потери функции (при правильном лечении), как правило, не наблюдается. Вовремя не устраненное нарушение в работе мышц приводит к травматизации хрящевых и капсульных структур, накоплению участков атрофий и зон износа связочного аппарата. Данные патологические изменения приводят к снижению функциональных возможностей звеньев скелета, что может значительно влиять на биомеханику позвоночника.
Наиболее удобно позвоночник рассматривать через разделение на субсистемы, обеспечивающие основные функции позвоночника. Разделение условно и используется только для упрощения восприятия.
Концепция Панджаби (Manohar Panjabi). Стабилизирующие системы позвоночника.
Стабилизация тела и позвоночника в пространстве относительно вектора гравитации зависит от слаженной работы трех взаимосвязанных и взаимозависимых субсистем.
Пассивная субсистема включает в себя позвонки, межпозвонковые суставы, межпозвонковые диски, связочный аппарат позвоночника, капсулы суставов. Также к пассивной части относят глубокие и поверхностные фасции и мышечные соединительнотканные оболочки.
Активная скелетно-мышечная субсистема состоит из мышц и сухожилий, прикрепляющихся как непосредственно к позвоночнику, так и к другим костным рычагам опорно-двигательной системы.
Невральная субсистема или субсистема обратной связи состоит из рецепторов, информирующих ЦНС о параметрах силы, угловых ускорениях, силе и скорости растяжения/сокращения мышц и многое другое. Данные рецепторы располагаются в мышцах, связках, сухожилиях, капсулах суставов, фасциях.
Все три субсистемы работают синхронно, обеспечивая статическую и динамическую стабилизацию тела в пространстве и позвоночника в частности.
Поскольку человек постоянно находится в состоянии контролируемого неравновесия, то для каждого сустава и межпозвонкового диска существует точка (центр реакции), где суммируются моменты сил гравитации, мышечной тяги, груза и т. д. Невральная контролирующая субсистема отслеживает все параметры с механорецепторов, расположенных в связочной ткани и проприоцепторов, расположенных в мышечной ткани, с целью равномерного распределения нагрузки на отдельные суставы и максимально их минимизирует.
Всегда нужно помнить, что конструкция «модель человека – маятник» прежде всего костно-связочно-фасциальная, в которой сократительный компонент мышц играет второстепенную роль исполнителя или корректора движений. Только часть движений выполняется за счет активации сократительного компонента мышц, в то время как другая часть выполняется как за счет внешних сил (гравитация, инерция), так и внутренних (упруго-эластические свойства соединительной ткани). Иногда мышцы только начинают движение, а гравитация и инерционные силы его выполняют. Мышцы в данной модели – модулятор и корректор положения и движения. Избыточная мышечная активация приводит к увеличению нагрузки на суставы, что может привести к быстрому износу опорных структур – дисков, менисков и гиалиновых хрящей суставов.
Для удобства объяснения причин боли, механизмы я разобью на три части, согласно стабилизирующим системам позвоночника: боль мышечная, фасциально-связочно-дисковая, и боль при поражении невральных структур.
Сначала стоит рассмотреть механизм развития неспецифической боли, вызванной нарушением мышечной работы
Чтобы понять причину мышечной боли, следует бегло вспомнить некоторые необходимые факты о мышцах:
Для нашей темы достаточно рассмотреть мышцы через 3-х компонентную усредненную модель.
Любая мышца состоит из сократимой, эластичной и сухожильной ткани.
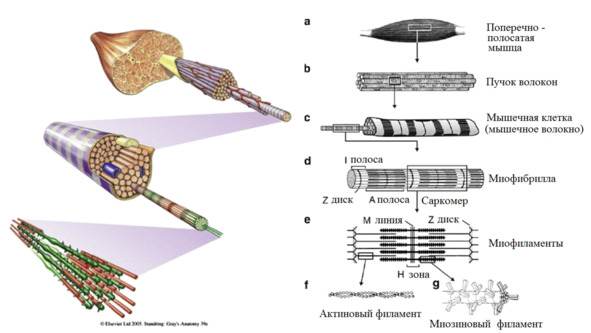
Мышечные сократительные волокна (мышечные клетки) подразделяют на два типа: экстрафузальные и интрафузальные.
Экстрафузальные – основной сократительный субстрат, вызывающий сокращение мышцы. Т.е. тот компонент, который обывателем воспринимается непосредственно как «мышца» или «мясо».

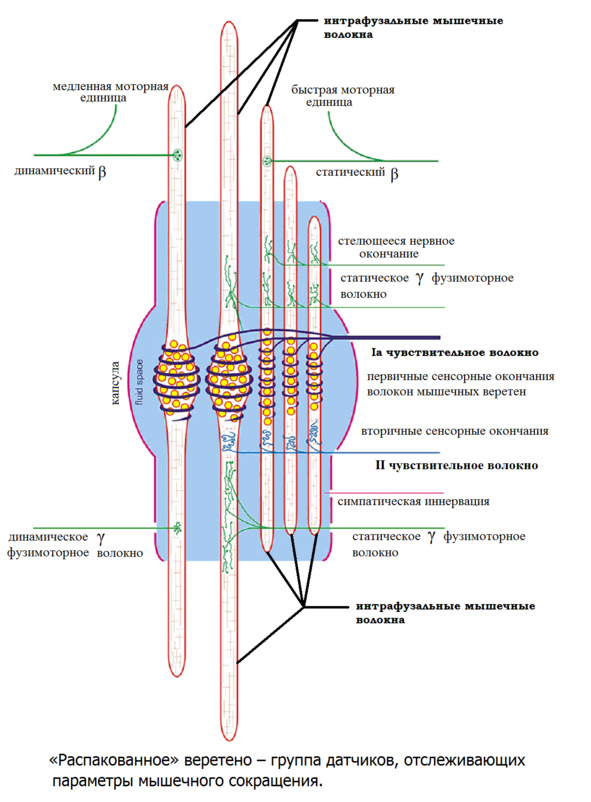
Интрафузальные (внутри веретена) – тонкие мышечные волокна, через напряжение/расслабление которых происходит настройка и регулировка чувствительности датчиков длины и скорости сокращения мышцы. Так называемое «веретено». Вклад в развитие силы вносят опосредовано, участвуя в реализации рефлекса растяжения (миотатический рефлекс).
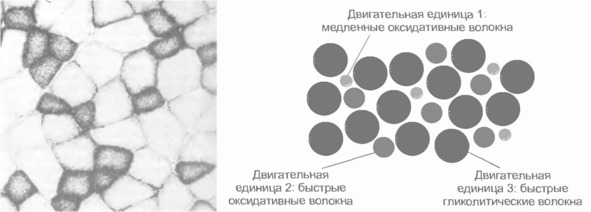
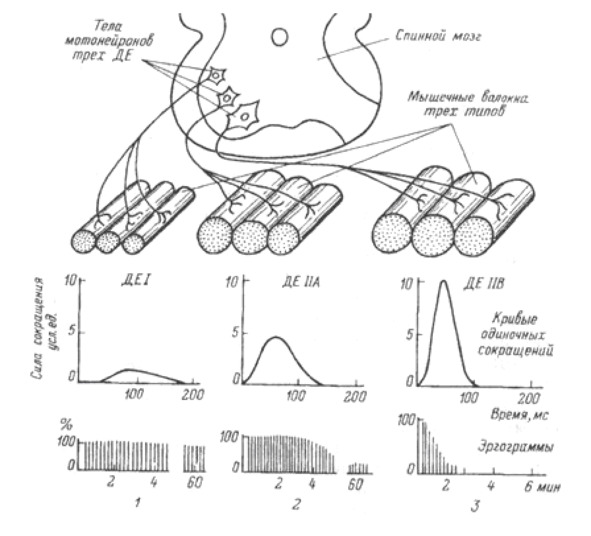
Экстрафузальные мышечные волокна подразделяются на три типа с разными характеристиками.
1. Медленные оксидативные волокна (I). Волокна I типа генерируют самую малую силу, но имеют наибольшую выносливость. Обеспечивают длительное сокращение мышцы; это используется для поддержания позы. Мембрана волокна не обладает электрической возбудимостью. Активация зависит от частоты стимуляции. Для данного типа волокна характерно медленное градуальное сокращение и медленное расслабление (приблизительно в 100 раз медленнее, чем у быстрых волокон).
Тип энергообеспечения – аэробное дыхание. Многие волокна при недостатке О2 продолжают работать за счет анаэробного гликолиза; в этом случае образуется лактат (молочная кислота) и создается кислородная задолженность.
2. Быстрые гликолитические волокна (IIb) Волокна II b обеспечивают самую большую силу, но обладают наименьшей выносливостью. Обеспечивают немедленное быстрое сокращение. Незаменимы при динамических нагрузках и скоростной локомоции. Мембрана волокна обладает электрической возбудимостью. Когда возникает активация волокна, развивается ответ типа «все или ничего». Мгновенное сокращение всех волокон в моторной единице. В качестве источника энергии интенсивно используется гликоген (анаэробный гликолиз, образуется лактат (молочная кислота). Сокращение в три раза быстрее, чем у медленных волокон. Довольно быстрое утомление.
3. Быстрые оксидативные волокна (IIa) Волокна IIa тип – универсальный. Обладают характеристиками как 1 так и 2 типа.

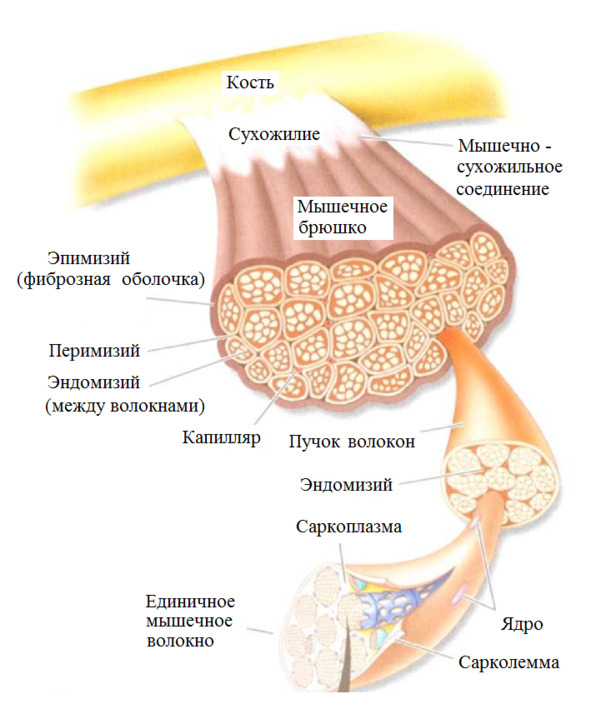
Каждое мышечное волокно окружено соединительной тканью (эндомизиумом). Пучки волокон также покрыты соединительно-тканной оболочкой (перимизиумом). Пучки объединены в целую мышцу с помощью эпимизиума. Вся мышца покрыта фасцией. Общая форма мышцы определяется соединительной тканью. Соединительная ткань преобладает над мышечными волокнами.
Эластические элементы подразделяют на последовательные и параллельные элементы. Последовательные эластические элементы (сухожилия) действуют по одной оси с сократительными элементами.
Параллельные эластические элементы (эндомизиум, перимизиум и эпимизиум) действуют параллельно оси действия сократительных элементов.
Каждая мышца состоит из трех типов мышечных волокон в разной пропорции. Зависит от глубины залегания и выполняемой функции. Чем «глубже» расположена мышца, тем больше в ней содержание медленных волокон. В поверхностных мышцах больше быстрых волокон. Мышечная композиция генетически предопределена.
Сила, которую мышца может развить благодаря своим эластическим элементам, будет максимальной, если мышца предварительно эксцентрически растянута внешней силой.
Сила, которую мышца может развить благодаря своим сократительным элементам, будет наибольшей, когда мышца начинает сокращение при длине, соответствующей состоянию покоя.
Общая сила, развиваемая совместно как эластическими, так и сократительными элементами, будет максимальной, если мышца предварительно эксцентрически растянута.
Эксцентрическая работа является работой сопротивления мышцы к ее растяжению внешней силой.
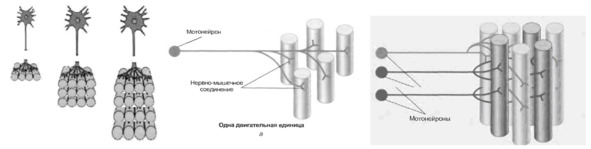
Управление сократительной активностью мышцы осуществляется с помощью большого числа мотонейронов – нервных клеток, тела которых лежат в спинном мозге, а длинные ответвления – аксоны в составе двигательного нерва подходят к мышце.
Система, включающая в себя альфа-мотонейрон, аксон и совокупность мышечных волокон, которые он иннервирует, получила название Моторная единица (МЕ) или двигательная единица (ДЕ).
Войдя в мышцу, аксон моторного нейрона разветвляется на множество веточек, каждая из которых подведена к отдельному волокну. Таким образом, один мотонейрон иннервирует целую группу волокон, которые сокращаются/расслабляются как единое целое.
Различают три типа моторных единиц.
Малый мотонейрон с малым количеством мышечных волокон. Имеет высокую степень возбудимости (низкий порог активации). Иннервирует медленные оксидативные волокна (кислород зависимые) (тип I).
Средний мотонейрон со средним количеством мышечных волокон. Имеет промежуточную степень возбудимости (средний порог активации). Иннервирует быстрые оксидативные волокна (тип II a).
Большой мотонейрон с большим количеством мышечных волокон. Имеет низкую возбудимость (высокий порог активации). Иннервирует быстрые гликолитические волокна (кислород независимые) (тип IIb).
Мышечные волокна отдельной моторной единицы всегда одного типа.
Мышца сокращается или расслабляется при взаимодействии двигательных и чувствительных нейронов.
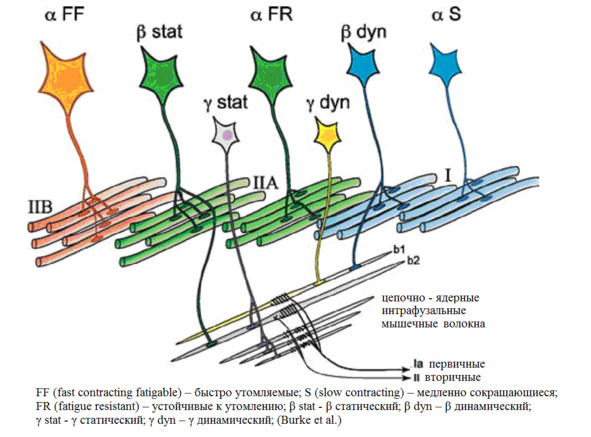
Схематично все «участники» мышечного сокращения/расслабления
(Manuel, M., & Zytnicki, D. (2011). Alpha, beta and gamma motoneurons: functional diversity in the motor system’s final pathway. Journal of integrative neuroscience, 10 (03), 243—276).
Альфа (α) -мотонейроны иннервируют экстрафузальные волокна. Гамма (γ) -мотонейроны – иннервируют интрафузальные. Бета (β) – одновременно иннервируют как экстра, – так и интрафузальные. Отвечают за слаженность сокращения обоих типов волокон, предотвращают потерю чувствительности «веретена» при сокращении экстрафузальных.
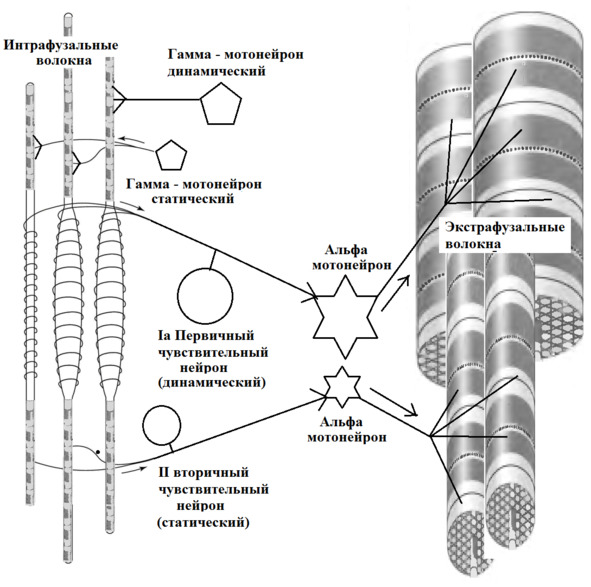
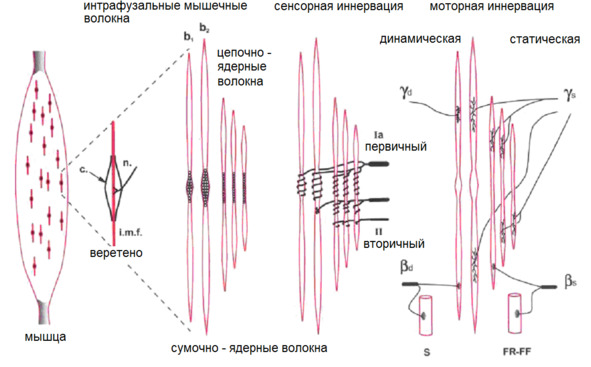
Схематично расположение «веретен» в мышце. В зависимости от специализации мышцы «веретена» могут иметь разное топографическое расположение среди мышечных волокон.
(Ellaway, P. H., Taylor, A., & Durbaba, R. (2015). Muscle spindle and fusimotor activity in locomotion. Journal of anatomy, 227 (2), 157—166).
В норме для развития максимальной силы мышцы должны активироваться все моторные единицы.
Функциональные свойства разных моторных единиц отражают особенности составляющих их мышечных волокон.
Совокупность мотонейронов, связанных с одной мышцей, называется пул мотонейронов. В нем представлены все три типа моторных единиц.
Синхронизация. При возбуждении одного альфа мотонейрона все иннервируемые им мышечные волокна одновременно сокращаются. Сила сокращения всех этих волокон будет суммироваться.
Последовательность активации моторных единиц называется рекрутинг.
Первыми активируются самые малые альфа-мотонейроны, связанные с медленными оксидативными волокнами.
Затем включаются быстрые оксидативные, чьи волокна крупнее. И наконец, активируются самые крупные мотонейроны, иннервирующие быстрые гликолитические волокна.
Сокращение медленных волокон может развивать до 15% максимальной силы мышцы.
Сокращение быстрых оксидативных может развивать до 45% максимальной силы.
Сокращение быстрых гликолитических волокон может обеспечивать максимальную силу сокращения.
Управлять силой мышечного сокращения можно только через усиление или ослабление частоты нервного импульса, вовлекая в работу новые моторные (двигательные) единицы, либо наоборот, выключая из работы задействованные МЕ.
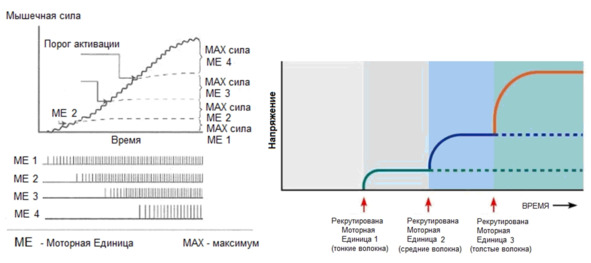
Каждая последующая по уровню порога активации моторная единица имеет большее количество мышечных волокон. Причем разница по количеству мышечных волокон между соседними моторными единицами растёт с увеличением порога возбудимости (активации) и если низкопроговые моторные единицы могут разниться в несколько мышечных волокон, то высокопороговые моторные единицы разнятся между собой уже в десятки или сотни мышечных волокон. Соответственно, активация таких волокон уже будет иметь не «дискретный» (градуированный) характер, а «взрывной».
У нетренированных людей количество одновременно сокращенных волокон не превышает 17%. В результате физических тренировок у высококвалифицированных спортсменов количество одномоментно активированных волокон составляет порядка 40%.
Исходя из этого можно сделать вывод, что наиболее точные и координированные движения можно совершать при наименьшем мышечном напряжении, когда, рекрутируя новые моторные единицы, ЦНС регулирует силу мышцы десятками новых мышечных волокон, вовлеченных в работу, а не тысячами.
Нарушение механизма последовательной активации/деактивации мышечных волокон серьезно влияет на всю систему управления движением.
Количество мышечных волокон, принадлежащих одной и той же моторной единице, увеличивается с возрастом. Это обусловлено изменением коэффициента иннервации, поскольку некоторые двигательные нейроны вырождаются и умирают, что приводит к тому, что аксоны других моторных единиц образуют большие ответвления и реиннервируют мышечные волокна, утратившие аксональную связь (происходит разрастание аксонов с захватом «чужих» мышечных волокон). Таким образом, с возрастом количество моторных единиц уменьшается, тогда как размер (количество мышечных волокон) тех моторных единиц, которые остаются, – увеличивается.
При постуральной (позной) деятельности в работу вовлекаются преимущественно низкопороговые медленные энергоэкономичные устойчивые к утомлению моторные единицы, а мышечные волокна, входящие в состав таких единиц, функционируют в режиме, близком к изометрическому и развивают при этом длительные тетанические сокращения небольшой силы, т.е. находятся в режиме постоянного сокращения.
В мышцах, для функции которых необходимы кратковременные сильные сокращения, больше быстрых волокон (фазные мышцы).
Рекрутинг – механизм последовательного включения сократительного аппарата мышцы, напрямую зависящий от выполнения поставленной на текущий период двигательной задачи (выдать необходимую силу, мощность, скорость, выносливость и т. д. то есть, выполнить необходимую работу с заданными параметрами). На основе механизма рекрутинга строится механизм межмышечной и внутримышечной координации. Другими словами, для сложных двигательных актов ЦНС должна координировать не просто схему включения/выключения отдельных мышц и мышечных групп, но и последовательность включения/выключения отдельных групп волокон внутри самой мышцы. Т.е., точно координировать и регулировать работу всех мышечных пулов в каждой отдельно взятой мышце. И главное в данном процессе является выключение мышечных волокон и возможность мышцы расслабиться и быть растянутой до первоначальной или заданной программой длинны. Из физиологии мышечного сокращения известно, что утомление не только снижает сократительную способность мышцы (соответственно вызывая потерю в силе), но и способность вовремя расслабляться, что увеличивает погрешности при управлении движением.
Чем выше процентное соотношение медленно сокращающихся тонических волокон, тем выше способность мышцы к накоплению и отдаче эластической энергии [рекуперация]. Соответственно тем выше экономичность самого движения.
КПД мышц достигает 35%, что является хорошим показателем производительности по сравнению с другими типами «двигателей», но даже при таком условии расход метаболической энергии на выполнение мышечного сокращения очень высок. И требуется значительное количество времени на пополнение энергетического запаса и накопления резервов.
Любое движение может быть представлено как перемещение из одной равновесной точки, в которой мышечная активность минимальна, в другую.
С точки зрения биомеханики поддержание равновесия и одновременное управление движениями являются достаточно сложными задачами, поскольку тело человека представляет собой многозвенную биомеханическую систему, в которой наиболее массивное звено (корпус) расположено высоко над площадью опоры, размеры которой ограничены площадью опорной базы.
Между звеньями существует сложное динамическое взаимодействие. Движение в любом отдельно взятом звене приводит к необходимости вырабатывать корректирующие силовые моменты (мышечное напряжение) во всех суставах кинематической цепи.
Для реализации любой двигательной задачи в рычажно-маятниковом механизме ЦНС нужно преодолеть проблему степеней свободы – в каждом определенном отрезке времени и траектории движения сегмента обеспечить его стабильность или мобильность относительно другого подвижного или неподвижного сегмента или опоры. Фактор длительности и силы напряжения мышц играют основную роль.
Ключевой функцией ЦНС является контроль избыточного движения, избыточного напряжения, стремление к минимизации степеней свободы или ограничение числа занятых независимых элементов движения.
Другими словами: ЦНС должна вовремя замыкать и размыкать суставы и контролировать скорость и углы сгибания-разгибания. При этом общий центр масс (ОЦМ) должен постоянно находиться под контролем ЦНС с целью препятствования падению.
Каждая мышца имеет свою индивидуальную структуру и специализацию. Но, несмотря на это, любое движение выполняется группами мышц посредством ансамблевого взаимодействия как внутри групп мышц со сходным действием, так и согласованно с группами антагонистов – мышц с противоположным вектором действия. Антагонизм довольно условен. При выполнении движения антагонисты выполняют контролирующую, тормозящую или стабилизирующую функцию по отношению к агонистам – мышцам, выполняющим основное движение.
Для выполнения различных задач ЦНС группирует мышцы в динамические мышечные цепи. Такие временные «союзы» направлены для решения конкретной задачи и не являются неизменными анатомическими образованиями. Такое групповое взаимодействие может требоваться для динамической стабилизации туловища (конечностей) или выполнения задач, требующих сложной координации. Одни и те же мышцы могут входить в состав различных мышечных цепей. Участие отдельных мышечных цепей различно даже при выполнении отдельных фаз движения. Чаще всего проблемы возникают при движениях со сложной структурой, когда одна и та же мышечная группа или отдельная мышца работает как агонист – выполняет основное движение, и стабилизирует сустав (одновременно являясь неотъемлемой частью системы стабилизации). Пример – поясничная мышца при сгибании позвоночника передними волокнами сгибает корпус, а задними контролирует углы сгибания позвонков относительно друг друга – стабилизирует поясничный отдел позвоночника – препятствует сгибанию корпуса.
Контроль за фазами перехода концентрического сокращения в эксцентрическое и обратно требует повышенного внимания со стороны ЦНС и точного контроля над отдельными мышечными параметрами – скоростью, силой сокращения, временем расслабления. Такая работа ЦНС обозначается как внутримышечная и межмышечная координация. Для предотвращения перегрузки и истощения отдельных мышц и для оптимизации выполнения и повышения точности движения ЦНС использует синергии – содружественную работу мышц, повышающую их производительность с одновременным снижением рабочего износа и повышением общего коэффициента полезного действия мышечной группы.
При любом движении мышца может работать как агонист, антагонист, синергист. Смена фаз зависит от выполняемого движения. Причем фазы могут сменяться мгновенно.
ЦНС применяет синергии для обеспечения гибкости и вариабельности выполнения двигательной задачи. Также такой механизм позволяет предотвратить или снизить истощение метаболических резервов в каждой отдельно взятой мышце.
Наряду с управлением синергиями, ЦНС использует и другой механизм мышечной работы – коактивацию – одновременное напряжение агонистов-антагонистов – мышц с противоположным действием. Используется для динамической стабилизации суставов.
В норме одновременное напряжение мышц (коактивация) не превышает 2—3 (два-три) процента от максимального мышечного сокращения. Правомерно будет сказать, что благодаря конструкции скелета и свойствам мышечно-фасциальной системы передвижение осуществляется почти в холостом режиме благодаря упруго-эластическим свойствам мышц и возможности работать в режиме рекуперации энергии.
При маятниковом механизме, активация мышечного сокращения выполняет корригирующую функцию по возвращению ОЦМ в площадь опоры или используется при средних и интенсивных нагрузках непродолжительный период. Все остальное время мышечная система работает в фоновом тоническом режиме с кратковременными пиками повышения активности различных мышечных групп.
Причины мышечной боли
Разбор начну с боли, вызванной спазмом мышц, поскольку данный механизм заявлен как основной и этот «концепт» прочно вбит в головы терапевтам, семейным врачам, массажистам и мануальщикам. В последнее время в литературе для неврологов навязывается такой же примитивный взгляд на боли в спине, как и у терапевтов. Если и дальше так пойдет, то по уровню знаний отличить невролога от семейного врача будет невозможно. Привожу конкретный пример:
Практическая неврология: руководство для врачей под редакцией профессора А. С. Кадыкова (Серия «Библиотека врача-специалиста), 2011. Предназначено для врачей-неврологов, врачей общей практики, терапевтов, фельдшеров, студентов медицинских вузов, а также для всех специалистов, оказывающих помощь больным с заболеванием нервной системы.
Глава 11. Боли в спине. Авторы: Л. С. Манвелов, В. В. Шведков, А. С. Кадыков):
«Первоначально основной причиной возникновения болей в спине считали воспаление нервных корешков и стволов. В дальнейшем инфекционно-аллергическая теория патогенеза болей в спине постепенно стала уступать место вертеброгенной, чему в определенной степени способствовал успех операций по поводу грыжи диска. Одно время все боли в спине стали объяснять дегенеративно-дистрофическими изменениями позвоночника, сдавлением грыжей межпозвонкового диска нервного корешка. В этот период появляется и соответствующая терминология: дискогенный корешковый компрессионный синдром, вертеброгенная радикулопатия, вертеброгенный рефлекторный синдром.
В 1980—1990—е гг. у неврологов стала превалировать теория преимущественно мышечного происхождения болей в спине. В настоящее время считается, что почти 90% случаев причиной болей в спине являются миофасциальные синдромы, [выделено мной] а на долю вертеброгенных нарушений приходится не более 10%. Это отражает и соответствующая терминология (дорсалгия, люмбалгия, миофасциальный синдром).


