


Полная версия
Математические модели в естественнонаучном образовании. Том II

Денис Соломатин
Математические модели в естественнонаучном образовании. Том II
Глава 5. Построение филогенетических деревьев
Смоделировав эволюцию ДНК в предыдущей главе, теперь готовы использовать эти модели для важных выводов из реальных данных ДНК. Увидим, как модель молекулярной эволюции вместе с некоторыми новыми математическими методами может быть использована для восстановления хронологии событий эволюционной истории. Давайте рассмотрим хорошо изученный, но все же удивительный вопрос: кем приходятся люди современным обезьянам? Более точно, какие из горилл, шимпанзе, орангутанов и гиббонов являются нашими ближайшими эволюционными родственниками, или все эти обезьяны более тесно связаны друг с другом, чем с нами?

Рисунок 5.1. Две возможные филогении гоминоидов.
Ранние эволюционисты рассматривали шимпанзе и горилл как наших ближайших родственников. Считалось, что люди и эти африканские обезьяны образуют одну эволюционную группу, которая отделилась от других линий обезьян в более отдаленном прошлом. Чуть позже господствующим стало мнение, что все современные человекообразные обезьяны более тесно связаны друг с другом, чем с людьми. Две возможные схемы, представляющие более подробные версии этих конкурирующих взглядов на эволюцию гоминоидов, показаны на рисунке 5.1.
Вопросы для самопроверки:
– Поскольку шимпанзе и горилла являются африканцами, в то время как орангутанг и гиббон являются азиатами, что, если вообще что-либо, каждое из этих деревьев укажет о вероятном месте появления первых людей?
Как выбрать, какое из этих или многих других возможных эволюционных деревьев является лучшим описанием происхождения гоминоидов? Один из подходов заключается в том, что сначала выбирают конкретный ген, общий для всех человекообразных обезьян и людей, но последовательность ДНК которого показывает вариации от вида к виду. Если предположить, что этот ген является общим и сходным для всех гоминоидов, поскольку он произошел от общего предка (то есть последовательности ортологичны, другими словами, к их разделению привел процесс видообразования), то вариации последовательностей среди видов должны содержать информацию об их эволюционной истории.
Например, 898-элементные пары последовательностей митохондриальной ДНК HindIII из этих гоминоидов и семи других приматов были зарегистрированы в работе Хаясака с соавторами 1988 года, он в свою очередь опирался на работу коллективов под руководством Андерсона 1981 года и Брауна 1982 года. Эти последовательности согласуются на 67-97% участков, в зависимости от того, какие из них сравниваются. Чтобы увидеть эти последовательности, загрузите базу данных primatedata в MATLAB, а затем посмотрите имена переменных, в которых они хранятся.
Хотелось бы вывести филогенетическое дерево, подобное одному из представленных на рисунке 5.1, показывающее, как все обезьяны эволюционировали от общего предка. Но какие данные указывают на «лучшее» дерево или даже «хорошее» дерево, чтобы объективно описать эволюционное происхождение?
Конечно, ученые уже рисовали деревья, показывающие подозрительные эволюционные отношения между видами задолго до появления методов секвенирования ДНК. Морфологическое сходство между видами является одним из источников подсказок относительно того, какие деревья адекватно описывают происхождение видов. Идентификация общих предков по окаменелостям – это другой подход. Теперь данные последовательности предоставляют новый источник информации об эволюционной истории, но использование их для вывода филогенетических деревьев требует разработки новых математических инструментов.
5.1. Филогенетические деревья
Прежде чем начнем разрабатывать методы построения филогенетических деревьев, понадобится некоторая терминология. Поскольку последовательности, которые, возможно, захотим связать, могут происходить от разных видов, как в примере с гоминоидами, или вместо этого от разных подвидов, популяций или даже отдельных особей, будем называть каждый источник последовательности ДНК таксоном (множественное число таксоны). Эквивалентный общеупотребительный термин – операционная таксономическая единица, обычно обозначаемая аббревиатурой ОТЕ (в иностранной литературе можно встретить обозначение OTU).
Будет стараться нарисовать диаграмму, состоящую из отрезков линий, которая представляет собой эволюционную историю таксонов. Каждый из сегментов линии на диаграмме по устоявшейся в теории графов терминологии называется ребром. Диаграмма, подобная приведенной выше, в которой нет циклов и петель, образованных ребрами, называется деревом.
Вопросы для самопроверки:
– Почему разумно предположить, что эволюционные отношения могут быть смоделированы путем рисования именно деревьев? Что бы это значило, если бы существовали цикл или петля?
Поскольку существует боковой перенос генов, например, когда вирусная ДНК постоянно включается в ДНК хозяина, деревья не могут описать все эволюционные отношения. Они обеспечивают простейшую модель, которая, тем не менее, полностью адекватна для большинства применений.
Точка, в которой сходятся нескольких ребер называется внутренней вершиной, в то время как висячий конец ребра у таксона называется конечной вершиной или листом дерева. Вершина, в которой будет находиться общий предок всех таксонов, называется корнем.
Говорят, что дерево раздваивается, находится в состоянии бифуркации, если на каждой его внутренней вершине встречаются по три ребра, а у корня сходится два ребра, как на деревьях на рисунке 5.1. Такие деревья называют двоичными или бинарными. Хотя с биологической точки зрения возможно, что дерево, отличное от двоичного, могло бы описывать эволюционную историю, обычно эту возможность игнорируют.
Вопросы для самопроверки:
– Каково было бы эволюционное значение вершины в дереве, где встречаются четыре ребра (то есть, где маршрут из одного ребра расходится на три направления)? Можете ли представить себе правдоподобные обстоятельства, при которых несколько видов могут расходиться таким образом?
Хотя в идеале каждое филогенетическое дерево должно иметь корень, показывающий общего предка таксонов, иногда приходится обходиться без него. Некоторые методы филогенетического построения деревьев дают некорневые деревья. Например, на рисунке 5.2 показано некорневое дерево и несколько корневых деревьев, которые с ним согласуются. Два дерева справа могли быть согнуты и растянуты, чтобы выглядеть как дерево слева; их отличает только расположение корня.

Рисунок 5.2. Некорневое дерево (слева) и две его корневые версии (в центра и справа).
Посмотрим на деревья с топологической точки зрения. Дерево, относящееся к ряду таксонов, может фактически указывать несколько различных типов информации об их отношениях. Во-первых, если не указываем длины ребер, а значит, смотрим только на ветвящуюся структуру, то рассматриваем только топологию дерева. Считается, что два дерева топологически одинаковы, если можно согнуть и растянуть ребра одного из них, чтобы получить второе дерево. Однако нельзя отрезать ребро и снова прикреплять его в другом месте; это может дать дерево, которое топологически отличается от исходного.
На рисунке 5.3 деревья




Для корневых деревьев используем аналогичную концепцию. Два корневых дерева топологически эквивалентны, если одно можно преобразовать в другое, не перемещая корень. Можно изменить длину ребер, но не структуру ветвления.

Рисунок 5.3. Четыре топологических дерева; как некорневые деревья, все, кроме правого нижнего, они идентичны.
Вопросы для самопроверки:
– Как на рисунке 5.3 расположить корень дерева
Топологическое дерево, даже некорневое, довольно многое говорит об эволюционной истории таксонов, к которым оно относится. Например, все деревья на рисунке 5.2 показывают, что таксоны



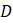
Знание местоположения корня передает больше информации и может дать лучшее представление о порядке событий во времени. Например, изображенное справа на рисунке 5.2 дерево однозначно задаёт следующий порядок бифуркаций: общий предок дал начало двум таксонам, один из которых, возможно, эволюционировал дальше, чтобы стать

Дерево в центре рисунка 5.2 можно интерпретировать аналогичным образом. Общий предок дал начало двум таксонам, один из которых дал начало как
Количество различных топологических деревьев, которые могут соотносить несколько терминальных таксонов, быстро растёт с увеличением числа таксонов. Например, существует только 1 некорневое топологическое дерево, относящееся к 3 таксонам, но есть 3 некорневых топологически различных дерева, относящиеся к 4 таксонам.
Вопросы для самопроверки:
– Нарисуйте одно некорневое топологическое дерево, которое может относиться к терминальным таксонам
На 5 терминальных таксонов приходится 15 таких деревьев. Таким образом, если не принимать во внимание местонахождение корня, существует на 13 деревьев, которые могут связать 5 гоминоидов, а больше, чем было представлено во введении к главе. Для 6 терминальных таксонов насчитывается более 100 возможных некорневых деревьев. По мере увеличения числа таксонов количество деревьев быстро вырастает до астрономических размеров. В упражнениях найдете точные формулы, определяющие количество некорневых и корневых деревьев, относящихся к
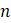
На помощь в решении обозначенной проблемы поиска лучшей классификации приходят метрические деревья. В дополнение к топологической структуре дерево может иметь метрическую структуру; каждому ребру может быть присвоена определенная длина. Эта метрическая структура может быть задана путем записи чисел для обозначения длин рядом с ребрами (см. Рисунок 5.4 (слева)), или ребро может быть наглядно представлено путем рисования дерева с ребрами соответствующей длины, но без их явной нумерации. Таким образом, топологическое дерево и немаркированное метрическое дерево неотличимы друг от друга. Для ясности, будем маркировать ребра их длиной, когда нужно задать метрическое дерево.
Как правило, длины ребер в филогенетическом дереве, построенном из данных последовательности ДНК, каким-то образом представляют собой количество мутаций, которые произошли между расщеплениями линии. Чем длиннее ребро, тем больше последовательность ДНК мутировала в ходе эволюции, которую представляет это ребро.
Если, например, модель Джукса-Кантора замещения оснований адекватно описала эволюцию нескольких таксонов, то длина ребра в дереве, относящемся к ним, может быть расстоянием Джукса-Кантора между последовательностями на двух концах. Как видели в главе 4, это расстояние представляет собой среднее число замен оснований на сайт, произошедших при происхождении новой последовательности. Сюда включены мутации, скрытые другими мутациями, для оценки которых была разработана формула расстояния. Поскольку расстояние Джукса-Кантора является аддитивным и симметричным, общее расстояние между двумя таксонами вдоль дерева должно быть расстоянием Джукса-Кантора между ними.
Если предположение о молекулярных часах справедливо для эволюции связанных последовательностей, то расстояния в дереве имеют постоянное значение. Напомним, что молекулярные часы просто означают, что скорость мутаций постоянна для всех рассматриваемых линий. Если


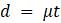
Таким образом, молекулярные часы означают, что количество мутаций на любом ребре пропорционально прошедшему времени, при этом константа пропорциональности представляет собой постоянную скорость мутации. Если предположить, что существуют молекулярные часы, то независимо от того, рисуем ли длины ребер, представляющие количество мутаций или только прошедшее время, то нарисуем одну и ту же фигуру с точностью до масштаба этой константы.
Если гипотеза молекулярных часов справедлива для корневого метрического дерева, то каждый лист будет расположен на одинаковом общем расстоянии от корня дерева. Это связано с тем, что расстояния от корня пропорциональны времени, прошедшему с тех пор, как таксоны начали расходиться с общим предком. У каждого таксона было одинаковое количество времени, чтобы эволюционировать от корневого предка, поэтому каждый таксон накопит одинаковое количество мутаций.
Без молекулярных часов связь между количеством мутаций вдоль ребра и количеством времени может быть сложной для моделирования. Предположим, что вдоль одного ребра филогенетического дерева частота мутаций была довольно мала, а вдоль другого – частота мутаций была большой. Затем несмотря на то, что оба края могут соответствовать одинаковому количеству времени, вдоль одного из них произойдет значительно больше мутаций. Без получения какой-либо дополнительной информации о скорости мутации – возможно, путем сравнения с летописью окаменелостей – обычно нет способов определения прошедшего времени, связанного с ребрами деревьев.
Метрические деревья иногда рисуются в «квадратном» стиле, чтобы было легче сравнивать расстояния по различным эволюционным путям. Например, два дерева на рисунке 5.4 представляют одну и ту же информацию. В дереве слева ребра имеют указанную длину, а в дереве справа горизонтальные ребра имеют те же длины. Таким образом, вертикальные ребра на правом дереве считываются как не вносящие никакого вклада в количество мутаций; они служат исключительно для разделения различных линий для повышения читабельности.

Рисунок 5.4. Разные изображения одного и того же метрического дерева.
Задачи для самостоятельного решения:
5.1.1. Рассмотрим деревья на рисунке 5.5.

Рисунок 5.5. Деревья для задачи 5.1.1.
а. Какие из них совпадают с корневыми метрическими деревьями?
б. Какие из них совпадают с некорневыми метрическими деревьями?
в. Какие из них такие же, как корневые топологические деревья?
г. Какие из них такие же, как некорневые топологические деревья?
д. Для каких деревьев работают молекулярные часы?
5.1.2. а. Нарисуйте единственное топологически уникальное некорневое раздвоенное дерево, которое могло бы описать связь между 3 таксонами.
б. Нарисуйте три топологически различных корневых раздвоенных дерева, которые могли бы описать связь между 3 таксонами.
5.1.3. а. Нарисуйте все 3 топологически различных некорневых раздвоенных деревьев, которые могли бы описать связь между 4 таксонами.
б. Нарисуйте все 15 топологически различных корневых раздвоенных деревьев, которые могли бы описать связь между 4 таксонами.
5.1.4. Для


5.1.5. Для

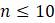
5.1.6. В этой задаче рассмотрим рассуждения, лежащие в основе формул для числа топологически различных деревьев, корневых и некорневых.
а. Предположим известно, что некорневое дерево с
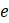
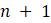

б. Поскольку некорневое дерево с 2 концевыми вершинами имеет 1 ребро, объясните из пункта (а), почему некорневое дерево с

в. Предположим известно, что существует


г. Поскольку существует только 1 некорневое дерево с 2 концевыми вершинами, объясните используя пункт (c), почему существует
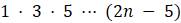

д. Объясните, почему
е. Почему число корневых деревьев с
ж. Сделайте вывод о правильности формул в задачах 5.1.4 и 5.1.5.
5.1.7. Поскольку митохондриальная ДНК у человека наследуется исключительно от матери, она может быть использована для структуры, относящейся к любому количеству людей из разных этнических групп, предполагая, что все люди произошли от одной первой человеческой самки. В зависимости от модели кластеризации этнических групп, это может дать представление о физическом местоположении той женщины, которую иногда называют митохондриальной Евой.
В работе Канна 1987 года была впервые предпринята попытка определить местонахождение митохондриальной Евы в Африке. Поддерживая теорию происхождения человека «из Африки», было построено дерево с корнями, которое, как утверждается, показывает отношения между 147 людьми. Сколько топологически различных деревьев нужно было бы рассмотреть, если бы действительно рассматривалась каждая возможность? Возможно, для ответа на этот вопрос придется использовать формулу Стирлинга:

5.1.8. Филогенез четырех терминальных таксонов A, B, C и D связан по определенному метрическому дереву. Суммарные расстояния между таксонами вдоль дерева оказались такими же, как в таблице 5.1.
Таблица 5.1. Расстояния между таксонами для задачи 5.1.8
A B C D
A .6 .6 .2
B .4 .6
C .6
а. Используя любой подход, который пожелаете, определите правильное некорневое дерево, относящееся к этим таксонам, а также все длины его ребер. Объясните, как исключили другие топологические деревья.
б. Можете ли определить корень дерева по этим данным? Объясните, почему да или почему нет.







