

 полная версия
полная версияИиссиидиология. Основы. Том 6
Глава 1. Биохимическая активность нейронных структур организма – основа амплиатизации Фокусной Динамики в ллууввумическом Направлении развития
6.1. Способность каждого нормально развитого (то есть не отягощённого патологическими физиологическими проблемами) человека консуетно осуществлять в своей системе Восприятия субтеррансивное проявление своеобразного квантового эффекта, который мы субъективно определяем как «личностное» Самосознание (или состояние внутреннего диалога с «самим собой», проявление которого при некоторых органических поражениях мозга становится невозможным), является, пожалуй, самым главным эгллеролифтивным Механизмом, обеспечивающим непрерывную и осознанную реализацию нашего психоментального творчества в окружающем нас Мире. Обширный и многообразный жизненный Опыт, приобретаемый при этом, позволяет каждому из нас всесторонне осознавать и персонифицировано отличать свойственные только «лично» нам визуальные, творческие и поведенческие особенности существования от подобных и других психоментальных проявлений, которые также характерны всему множеству остальных людей.
6.2. Наличие некоторой качественной разницы между энергоинформационными параметрами каждого ФПВ, образующейся в результате симультанного осуществления трансфинитного множества квантовых «смещений» внутри разнообразных паттернов Энерго-Информации, обусловливает параллельное проявление в нашей системе Восприятия другого квантового эффекта – нашей субтеррансивной Фокусной Динамики (ФД), которая, в свою очередь, качественно обусловлена наличием в фокусируемой нами НУУ-ВВУ-Конфигурации симультанной (интегральной, суммарной) резонационной активности разнокачественных состояний ФПВ Формо-Творцов бесчисленного множества других – молекулярных и клеточных (геномных, гормональных, нейротрансмиттерных и других) – Форм Самосознаний, структурирующих разные участки нашего биологического организма и по-разному влияющих на нашу общую ФД. Каждому человеку, как и любой другой Форме Самосознания, присуща своя собственная, субтеррансивная ФД.
6.3. Благодаря присутствию в наших системах Восприятия этого универсального эгллеролифтивного Механизма (выполняющего, кроме всего прочего, также функции адекватного сопоставления, анализа и субтеррансивного распознавания воспринимаемых нами вибрационных проявлений – форм, звуковых и визуальных явлений, ситуаций и отношений, и так далее), каждый из нас (как НУУ-ВВУ-Формо-Тип, одно из бесчисленных проявлений «личностных» Интерпретаций, симультанно структурирующих сллоогрентную ф-Конфигурацию нашей Стерео-Формы) имеет возможность качественно разотождествляться с Фокусными Динамиками всех остальных Форм Самосознаний (то есть осмысленно отделять «самих себя» не только от других людей, но также от животных, растений, природных явлений и всех прочих проявлений окружающей среды), что позволяет нам подобающим образом (субъективно, по-своему) отражать в своих поступках и психоментальных переживаниях профективную динамику окружающей нас действительности и более или менее эффективно регулировать свои отношения с представляющими её Формами Самосознаний.
6.4. Набор же уникальных Представлений «личностного» Самосознания, непрерывно генерируемых нашей Фокусной Динамикой благодаря наличию у нейронных Творцов нашего мозга консуетной резонационной связи со сллоогрентными Формо-Образами ноовременного «Континуума» (НВК), позволяет нам делиться собственными знанием и Опытом (посредством речи, письма, музыки, художественных произведений, научных открытий, технических устройств и так далее) с другими людьми, тем самым постоянно – хотя зачастую и неосознанно – пополняя оригинальными СФУУРММ-Формами общий амплификационный базис, обеспечивающий развитие всего Коллективного Сознания человечества.
6.5. Параллельно каждое из генерируемых нами Представлений особым образом клексуется и вибрационно маркируется (Творцами гиппокампа [рис. 7, Приложение], энторинальной коры [рис. 7, Приложение] и некоторых зон префронтальной коры [рис., п. 6.0354]) в нашей «субтеррансивной ОДС» – своеобразном вневременном «хранилище» всех сллоогрентных Формо-Образов нашей памяти и Опыта, отражающем характерные особенности инерционного осуществления нашего ротационного Цикла – условного периода нашей Жизни, начиная с момента активизации АСТТМАЙ-РАА-А-Парвулы в зачатке нашего мозга во время внутриутробного развития до очередного акта ревитализации, то есть биологической «Смерти» в данном «сценарии» развития. Формально ротационный Цикл для каждого из нас не прерывается никогда, так как субъективно и осознанно мы всегда способны воспринимать самих себя только живыми (будь то «внутриутробный», «межвозрастной» или «межформотипный» вид ревитализации), непрерывно и консуетно амицирируя (качественно «смещаясь») своей ФД из ф-Конфигураций одних «личностных» Интерпретаций в другие.
6.6. В свою очередь, проявление каждого из этих Представлений создаёт необходимые творческие возможности для симультанной реализации (в разных пространственно-временных режимах и исторических условиях!) разноуровневых (по типу вибраций) Формо-Творцов первой и – частично! – второй пар ИИССИИДИ-Центров, которые симультанно обеспечивают эксгиберацию трансфинитного множества самых разнообразных гетерогенеусных Форм Самосознаний, отчасти использующих в свойственных им режимах проявления Поля-Сознания (осмысленные паттерны Энерго-Информации, структурирующие СФУУРММ-Формы) генерируемых нашей системой Восприятия Представлений для организации и качественного преобразования собственных паттернов Энерго-Информации (как и мы с вами резонационно – и чаще всего неосознанно! – пользуемся в своём жизненном творчестве Полями-Сознаниями СФУУРММ-Форм, сформированных ими в своих группах ПВК).
6.7. Психоментальная основа самого «личностного» Самосознания (главной формой реализации которой служит наша Фокусная Динамика, обеспеченная симультанным функционированием систем Восприятия множества ротационно-резонационно сменяемых нами НУУ-ВВУ-Формо-Типов) консуетно и непрерывно (то есть вне зависимости от процесса постоянного осуществления нами ревитализационных актов – в виде бесчисленных, никак не осознаваемых нами, «смен-смертей» множества последовательно фокусируемых нами биологических тел) пополняется определённым субтеррансивным Опытом. Этот амплификационный набор наших разнокачественных субъективных Представлений симультанно синтезируется в разных «сценариях» нашего инерционного развития совместными реализационными усилиями Формо-Творцов различных Уровней нашего «коллективного Бессознательного» (инстинктов) и «коллективного Подсознания» (творческой Интуиции) – за счёт постоянной внутрибирвуляртной амплиатизации более простых и примитивных (разнопротоформных) Представлений во всё более сложные и глубокие (ллууввумические).
6.8. То есть этот амбигулярный инерционный процесс поквантового формирования и конкатенационной амплиатизации Фокусной Динамики нашего «личностного» Самосознания совместно и параллельно обеспечивается Формо-Творцами как более деплиативных («примитивных», дооллсово-лоолгсовых, представляющих «коллективное Бессознательное»), так и более совершенных и универсальных (димидиомиттенсных, транслюценсных, представляющих «коллективное Подсознание») – по отношению к ныне фокусируемым нами микстумным НУУ-ВВУ-Формо-Типам – Форм Самосознаний, которые в данных условиях эксгиберации отражают характерные творческие тенденции не только ллууввумических, но также и самых разных Коллективных Космических Разумов (Прото-Форм) и типов бирвуляртности.
6.9. Когда мы говорим о реализационных проявлениях в нашей Жизни тех или иных Уровней нашего «личностного» Самосознания, то прежде всего имеем в виду сложнейший Механизм по проявлению в нашей ФД Полей-Сознаний различных субъективных СФУУРММ-Форм, отражающих сочетания каких-то качественных признаков, которые свойственны тем или иным ОО-УУ-Сущностям и которые мы в состоянии синтезировать через амбигулярную деятельность Творцов нашей системы Восприятия. Упрощённо говоря, одна часть амплификационной функциональности этого Механизма, реализуемой через нашу ФД, обеспечена информационным эфирным содержимым сллоогрентных СЛУИ-СЛУУ-конструкций НВК (факторными «осями», АСТТМАЙ-РАА-А-Парвулами, ФАТТМА-НАА-А-Артикулами, виваксами, виталитасами и так далее), в то время как другая её часть обусловлена симультанной совокупной активностью Фокусных Динамик бесчисленных Творцов всевозможных разнопротоформных и разнокачественных Форм Самосознаний, плотно структурирующих системы Восприятия фокусируемых нами микстумных НУУ-ВВУ-Формо-Типов (каузальных «Каналов», ядерного генома, клеточных и всех остальных формо-структур нашего организма).
6.10. В основе функционирования нашего мозга лежат химические, электрические и другие (пока что неизвестной нам природы) волновые процессы, симультанно и по-разному осуществляемые Творцами множества разнообразных формо-структур каждой из около 90 млрд нервных клеток – нейронов [рис. 5, Приложение], энергоинформационно обеспечивающие функции Форм Самосознаний которых можно условно сравнить с принципами работы конденсатора, имеющего отрицательный заряд внутри и положительный – на поверхности. В спокойном состоянии между внутренней и внешней средой каждого нейрона существует разность потенциалов – мембранный потенциал (около 75 милливольт), который образуется в силу наличия на поверхности нейрона особых белковых молекул (рецепторов), выполняющих роль своеобразных натрий-калиевых насосов.
6.11. Их Творцы – за счёт энергии АТФ, выделяемой ими при гидролизе нуклеотида, – направляют ионы калия внутрь клетки, а ионы натрия – наружу, на поверхность нейрона, покрытую ветвящимися отростками – дендритами, которые являются основной воспринимающей формо-структурой каждого нейрона. В зависимости от расстояния до тела своего нейрона, дендриты делятся на близкие и удалённые (последние могут образовывать полуавтономно функционирующие модули). К каждому дендриту подходят аксонные окончания других нейронов, образуя в каждой из точек их совместного взаимодействия множество синапсов [рис. 12, Приложение] – специализированных межклеточных контактов, передающих возбуждения между Творцами разных нейронов (а также нейронов и мышечных, секреторных клеток).
6.12. Бóльшая часть синапсов формируется «вокруг» множества своеобразных мембранных выростов разного размера (так называемых «шипиков»), которые образуются на поверхности каждого дендрита и чьё назначение заключается в «смешивании» Информации, симультанно образующейся в синапсах разных нейронов, в результате чего она получает возможность характерным для неё образом симультанно влиять на творческие состояния их Творцов, тем самым совместно синхронизируя их функции в общем направлении реализации этой Информации. Чем более сложную функцию выполняет нейрон, тем больше различных анализаторов посылают ему необходимые сведения, и тем больше шипиков формируется на его дендритах (например, количество шипиков на пирамидных нейронах двигательной зоны коры головного мозга может достигать нескольких тысяч!). Когда какой-то из шипиков или целая группа их продолжительный период бездействуют, не получая свойственную им Информацию, то они исчезают с поверхности дендрита.
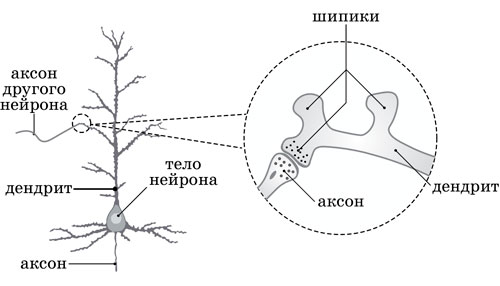
Рисунок 6.0012. Схема нейрона (дендрит с шипиками)
6.13. В силу того, что дендритные разветвления одних нейронов часто очень плотно переплетаются и тесно соприкасаются с подобными разветвлениями других нейронов, синапсы одного дендрита, сформированные на шипиках разной длины, могут располагаться гораздо ближе к мембране рядом находящегося дендрита (или дендритов других нейронов), чем даже его собственные синапсы, в результате чего между шипиками нескольких плотно прилегающих друг к другу нейронов может образоваться общий для них пространственно распределённый сигнал (сложнокомбинированный синапс), представляющий собой интеграл совместной синхронной активности Творцов, структурирующих разные нейротрансмиттеры. Это обстоятельство может сыграть весьма значимую роль в процессе передачи и запоминания Информации, осуществлённого через данную группу нейронов.
6.14. Благодаря такому – синаптическому – взаимодействию Творцы нейрона реагируют на поступающие к ним сигналы и способны сами при необходимости генерировать наружу собственный импульс (волну деполяризации или спайк – кратковременное резкое колебание, сопровождающее возбуждение в нервных или мышечных клетках). При этом нейронные Творцы реагируют на каждый паттерн Информации (стимул) не единичным импульсом, а целой серией – определённой волновой последовательностью, частота чередующихся импульсов в которой тем выше, чем точнее Творцы смогли идентифицировать характерный для данного нейрона Формо-Образ, проявляющийся в их ФД в виде определённого стимула (то есть чем выше степень их совместного резонирования с данной Информацией, тем выше частота передаваемых ими импульсов).
6.15. Кроме генерируемых импульсов, Информация между Творцами некоторых нейронов также может «проецироваться» за счёт наличия у них специальных совместных белковых канальцев, которые соединяют их внутриклеточное содержимое и позволяют транслировать электрические потенциалы напрямую, путём перехода ионов из одной клетки в другую и образования электрических синапсов (этот способ относительно редко встречаются в нашей ЦНС). Но помимо синаптических – химического и электрического – способов взаимодействия, между Творцами разных нейронов непрерывно осуществляется также и спонтанная (резонационная) волновая Творческая Активность, обусловленная симультанной инициацией ими в НВК Формо-Образов общего для них «сценария» развития.
6.16. Эта спонтанная волновая активность, инициируемая Творцами ДНК и поддерживаемая всеми прочими внутриклеточными Творцами, всегда фоново присутствует буквально в каждом нейроне (а также во всех зонах головного мозга живого человека), вне зависимости от процесса генерации им синаптических импульсов (например, если отключить весь мозг от поступления внешней Информации, то электромагнитный тип активности между нейронными Творцами сенсорных зон только повысится). К слову сказать, очень важную роль в создании и поддержании в нашем Самосознании этого волнового источника Энерго-Информации выполняют формо-структуры находящейся над мозгом паутинной оболочки (организована в виде тонкой «паутины» из соединительной ткани и глиальных клеток), которая, натянувшись над извилинами мозга, тем не менее не заходит ни в них, ни в борозды.
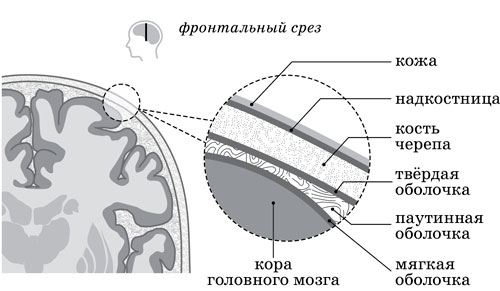
Рисунок 6.0016. Схема расположения оболочек головного мозга
6.17. Тоненькая, полупрозрачная и гладкая паутинная оболочка выполняет функции своеобразного рассеивателя электромагнитных волн миллиметрового диапазона (в пределах от 1 до 10 мм и частотой от 30 до 300 ГГц), симультанно генерируемых Творцами всех участков коры и подкорки. А вместе с уникальными особенностями формо-структур мягкой оболочки (прозрачная мембрана, покрывающая наружную, нижнюю и внутреннюю поверхности головного мозга; заходит во все щели и борозды, обильно структурирована крупными и мелкими сосудами) они представляют собой тончайший по своей функциональности механизм для воспроизведения (совместно с эпифизом) сллоогрентных СФУУРММ-Форм в нашей системе Восприятия (а также реализации всевозможных грёз, фантазий, «видений» и галлюцинаций в виде причудливой игры в нашем Воображении постоянно меняющихся Формо-Образов НВК). Здесь формо-структуры мягкой мозговой оболочки служат своеобразным голографическим «биоэкраном» для отражения Полей-Сознаний и СФУУРММ-Форм, совместно генерируемых в виде электромагнитных волн ядерными Творцами наиболее синхронизированных в данный момент участков мозга.
6.18. Как вы уже знаете, все внутриклеточные Творцы (включая и геномных) обеспечивают поддержание высоких уровней собственной самоорганизации и энергоинформационного обмена за счёт наличия у них очень важной особенности – способности симультанно пребывать в наших с вами нынешних режимах эксгиберации в специфическом состоянии квантовой нелокальности («спутанности»), осуществляя свою деятельность на условной «границе» между так называемыми «фермионным и бозе-эйнштейновским» видами плазмы (формируют всю энергоинформационную основу взаимосвязей внутри ПВК) и эфирными структурами ноовременного «Континуума».
6.19. Благодаря этому они, резонационно инициируя в НВК присущими им (их Фокусным Динамикам) качественными состояниями (волновыми генерациями) соответствующие Формо-Образы, постоянно обеспечивают самих себя всеми необходимыми «инструкциями поведения» в каждый условный момент реализации своей совместной Творческой Активности, «проецируя» получаемую ими Информацию не только внутри собственной клетки (для коллективной организации своих функций), но и далеко за её биологические пределы (в качестве обмена опытом с Творцами других клеток). Эти волны миллиметрового диапазона фоновой симультанной активности всего разнообразия Творцов нейронов распространяются как внутри любой из локальных зон коры, так и по всем остальным участкам мозга (через проекционные взаимосвязи), практически мгновенно (в нашем понимании!) перенося Информацию от одних частей нашего тела к остальным его частям и органам.
6.20. Проницая свойственной ей вибрацией какую-либо область коры, каждый, генерируемый нейроном, паттерн волн с закодированной в нём Информацией о каком-то конкретном событии или явлении, благодаря голографическим особенностям мягкой мозговой оболочки, всегда сохраняет свою уникальную волновую ф-Конфигурацию, что позволяет нейронным Творцам (через ядерный геном) любого участка коры всегда иметь возможность знать, что происходит в данный момент в других частях мозга и тела, и, помимо синаптических связей между собой, амбигулярно (учитывая обстоятельства) и спатиумально (дистанционно) координировать с Творцами других участков все свои функции и «текущие» реализационные усилия.
6.21. Объём этой фоновой Информации, симультанно генерируемой миллиардами внутриклеточных Форм Самосознаний, структурирующих десятки триллионов клеток нашего организма (не считая археи, грибы, вирусы и другие микроорганизмы), поистине грандиозен и упорядочивается между Фокусными Динамиками всего множества разнокачественных групп Творцов только благодаря всеобщему действию на них единого Принципа Резонационности, а также наличию у Пространства-Времени особых свойств (в том числе «фрактальности», сллоогрентности). Благодаря своим микроскопическим размерам и характерным особенностям своей ФД, Творцы ДНК и остальных клеточных структур субъективно осознают самих себя в совершенно иных, чем у нас с вами, режимах мерности, что и позволяет им более или менее успешно управляться со столь огромными и стремительно (для нас!) меняющимися энергоинформационными потоками. В их же собственных системах Восприятия все наши психоментальные реакции и действия осуществляются в большей степени медленно и продолжительно, чем то время, в течение которого мы их субъективно переживаем и осуществляем.
6.22. К слову сказать, недавно учёными было обнаружено явление танатотранскрипта, суть которого сводится к тому, что по истечении уже нескольких часов после биологической смерти мозга и полного прекращения работы сердечно-сосудистой системы в нейронах мозга умершего самопроизвольно активизируются (обычно на срок до 5 суток) определённые участки ядерного генома – так называемые «зомби-гены», конкретные функции и назначение которых пока ещё не выяснено. Но достоверно известно то, что активность данных генов можно наблюдать в ДНК нейронов не только после осуществления акта ревитализации (наблюдаемой нами «Смерти личности»), но также и в эмбриональный период онтогенеза (сразу же после рождения эти гены сами дезактивируются). Комплексное рассмотрение данного феномена даёт нам основание рассматривать его как одно из множества сопутствующих биологических проявлений эдревитализационного (околоревитализационного) процесса.
6.23. Что же касается химического способа передачи Информации в синапсах, то он осуществляется при наличии в терминали отростка аксона, находящегося рядом с дендритом (или с терминалью аксона другого нейрона), молекул тех или иных нейротрансмиттеров, которые выделяются через этот аксон и характерны для каждого из образуемых им синапсов. Один нейрон может сразу выделять тысячи молекул разных видов нейромедиаторов, Форма Самосознания каждого из которых обладает характерной только для неё вибрацией, на которую тут же резонационно активизируются Творцы какой-то из находящихся на мембране нейрона белковых молекул – рецепторов, которые непосредственно и взаимодействуют с нейромедиаторами или нейромодуляторами. Располагаясь в синаптической щели, рецепторы выступают в роли транспортных ионных каналов, по которым внутрь нейрона перемещается определённая порция ионов.
6.24. Вещество, которое взаимодействует только с определённым типом рецепторов, называют лигандом данного рецептора. Именно от характера их совместного взаимодействия зависит то, будет или не будет сгенерирован импульс (реакция на стимул) в результате сложения вибраций только что поступившей к нейрону Информации (внешних сигналов). Кстати, рядом располагающиеся на мембране нейрона рецепторы могут соединяться между собой и образовывать димеры, из которых формируются целые специализированные ансамбли рецепторов (рецептивные кластеры), чья структура способна отражать повышенную в данном участке мембраны чувствительность на поступление сигналов лишь какого-то определённого типа.
6.25. Каждый лиганд, обладая свойственной только ему вибрацией, побуждает к открытию канал резонационно соответствующего ему рецептора, в результате чего мембрана нейрона в этом месте либо деполяризуется, либо гиперполяризуется, – в зависимости от того, какие именно из рецепторных каналов активизировались и, соответственно, какой из типов синапсов при этом смог сформироваться: при возбуждающем воздействии открываются каналы, пропускающие внутрь нейрона катионы (мембрана деполяризуется, тем самым понижая заряд нейрона), а при торможении открываются каналы, проводящие анионы (мембрана гиперполяризуется, внутриклеточный потенциал возрастает).
6.26. Также следует иметь в виду, что через синапсы каждого нейрона сразу способно «проецироваться» множество сигналов, так как количество симультанно образуемых одним нейроном синапсов может исчисляться десятками тысяч, а минимальная порция одноразового выделения нейротрансмиттера в синаптическую щель составляет от 6-7 тысяч и более молекул. Причём в силу избыточного количества выделяемого медиатора, часть его молекул распространяется также и за пределы данной синаптической щели, распыляясь (хотя и с гораздо меньшей плотностью) в среде, образованной между телами нейронов и телами окутывающих их глиальных клеток [рис., п. 6.0225] (это явление называется спилловером).
6.27. В силу постоянного образования в синаптической щели подобных «переливов» в пространстве между рядом находящимися синапсами избытки распыляемых ими тормозных и активирующих нейротрансмиттеров создают определённую интерференционную картину (взаимоналожение волновых эффектов, ведущее к их взаимному усилению в одних точках пространства и к ослаблению в других), каждый раз вызывая активность в каком-то месте мембраны, в результате чего рецепторы могут краткосрочно входить либо в состояние большего выпячивания на поверхности мембраны (нейрон становится чувствительным к данной Информации и может отреагировать на её повторение генерацией нового импульса), либо в состояние углубления в неё (нейрон становится невосприимчивым к данной Информации). Это свойство рецепторов имеет важное значение в процессе формирования у нас краткосрочной, оперативной памяти.







