Полная версия
Эволюция. Классические идеи в свете новых открытий



Несколько фактов о сексе, которые должен знать каждый
Эукариоты и прокариоты занимаются сексом совершенно по-разному.
Эукариоты практикуют «половое размножение», или, по-научному, амфимиксис. Что бы ни подумал читатель, но это значит, что у них в жизненном цикле присутствует два особых события. Первое – редукционное (т. е. «уменьшающее») деление, или мейоз, при котором из клетки с удвоенным набором хромосом (диплоидной) получаются клетки с одинарным набором (гаплоидные). Клетки, прошедшие редукционное деление, либо сразу становятся половыми клетками (гаметами), как у животных, либо предварительно сколько-то раз делятся обычным способом (митозом) и только после этого дают начало гаметам (так обстоит дело у растений). Второе событие – оплодотворение, или сингамия. При оплодотворении две гаметы, обычно происходящие от разных организмов, сливаются в одну клетку с двойным набором хромосом – зиготу. Зигота снова становится диплоидной, из нее вырастает диплоидный организм, который в какой-то момент путем мейоза начинает производить гаплоидные клетки.
При половом размножении смешивание генов двух родительских организмов, во-первых, взаимное (двустороннее), во-вторых – полногеномное. Отец и мать передают каждой гамете целый гаплоидный геном в одном экземпляре, т. е. ровно по одной копии каждого своего гена. Зигота получает два гаплоидных генома, по одному от каждого из родителей. В ходе мейоза эти два генома перемешиваются, причем весьма тщательно, так что в итоге в каждую гамету попадает более или менее случайная смесь аллелей, полученных данным родительским организмом от своих отца и матери. Соответственно, зигота получит примерно в равных пропорциях аллели обоих своих дедушек и обеих бабушек.
У прокариот (бактерий и архей) все по-другому. Участники полового процесса у них называются не родителями, а донором и реципиентом. Донор – тот, кто отдает часть своих генов и ничего не получает, реципиент – воспреемник чужих генов. Реципиент обретает новый генетический набор и сам становится как бы собственным потомком (а также потомком донора). Эта клетка с новыми генами дает начало своей линии потомков, размножающихся делением и время от времени передающих гены другим особям. Так что обмен генами у прокариот, во-первых, односторонний (от донора к реципиенту), во-вторых, не полногеномный, а фрагментарный, так как передается не целый геном, а лишь отдельные, обычно небольшие фрагменты ДНК.
Существует три разновидности прокариотического секса.
1. Конъюгация, при которой донор активен, а реципиент относительно пассивен. Донор прикрепляется к реципиенту при помощи специального отростка – пилуса (мн. ч. – «пили») и впрыскивает в реципиента немного своей ДНК. Часто этим процессом «руководят» маленькие эгоистичные[37] кольцевые хромосомы – конъюгационные плазмиды. Именно в плазмиде находятся все гены, необходимые для успешной конъюгации, включая гены белков пилуса. Первым делом плазмида перекачивает в реципиента не абы что, а копию самой себя. Часто на этом все и заканчивается. Такие плазмиды ведут себя как настоящие паразиты, используя одних бактерий для того, чтобы попасть в других. Скорее всего, они-то и «изобрели» конъюгацию как средство достижения своих корыстных целей. Однако плазмида может вступить в симбиоз с клеткой-хозяином – ведь, когда он делится, плазмида размножается вместе с ним, поэтому ей выгодно нести в себе гены, полезные для хозяина. Например, часто именно конъюгационные плазмиды распространяют в популяциях бактерий гены устойчивости к антибиотикам. Иногда плазмиды встраиваются в кольцевую хромосому хозяина. В этом случае вместе с плазмидной ДНК реципиент может получить и часть геномной ДНК донора.
2. Вирусная трансдукция. Переходя из клетки в клетку, вирус может прихватить с собой кусочек ДНК прежнего хозяина и встроить его в геном следующего. При трансдукции и донор, и реципиент пассивны. Активен только вирус.
3. Трансформация, при которой донор пассивен (по правде сказать, чаще всего он просто мертв), а реципиент активен. Трансформация – это захват микробами молекул ДНК из окружающей среды. Захваченные фрагменты могут принадлежать мертвым, разрушенным клеткам, но не обязательно: некоторые микробы выделяют ДНК во внешнюю среду еще при жизни. Проглоченная молекула ДНК может быть использована просто в качестве пищи, но может и встроиться в геном реципиента.
Чужие гены встраиваются в геном реципиента двумя способами. Первый способ – «незаконная» рекомбинация. Например, новый фрагмент ДНК может вставиться в хромосому вдобавок к уже имеющимся там генам. Именно так микроб может получить полезный новый ген от неродственного микроба. Такие заимствования могут иметь важные эволюционные последствия, их активно изучают и именно их обычно имеют в виду, когда говорят о горизонтальном переносе генов. Второй способ (гораздо более распространенный, хотя и менее известный, потому что его трудно изучать) – гомологичная рекомбинация. Суть его в том, что фрагмент ДНК донора замещает собой похожий фрагмент ДНК в хромосоме реципиента. Говоря упрощенно, реципиент вырезает из своей хромосомы кусочек и заменяет его похожим кусочком чужой ДНК. Как правило, это происходит при обмене ДНК между близкими родственниками, чьи геномы мало отличаются друг от друга. Если участки ДНК различаются сильно, гомологичная рекомбинация между ними технически невозможна (в гомологичной рекомбинации задействован принцип комплементарности, но мы не будем вдаваться в молекулярные подробности)[38].
Механизм гомологичной рекомбинации – очень древний. Он наверняка был у Луки – последнего общего предка всего живого. Он должен был появиться задолго до Луки, еще на заре РНК-мира. Используется он не только для перетасовки похожих фрагментов ДНК, но и для более очевидной и повседневной задачи, а именно для починки (репарации) повреждений ДНК, таких как разрывы двойной спирали. При этом фрагмент ДНК, гомологичный разрушенному, используется в качестве «заплатки» (да простят нас молекулярные биологи за такое вульгарное упрощение сложного процесса!).
О горизонтальном переносе генов (ГПГ) мы говорили в книге «Рождение сложности». Эволюционная роль этого явления огромна: микробы и одноклеточные эукариоты приобрели множество нужных признаков, заимствуя чужие гены. Геном любой бактерии прямо-таки напичкан явно «неродными», но при этом крайне полезными генами. Один из примеров – появление у цианобактерий способности к кислородному фотосинтезу. Событие, перевернувшее мир, превратившее Землю из унылой бескислородой «планеты микробов» в царство разнообразных, удивительных и сложных «высших» форм жизни. Как оно произошло? Исключительно благодаря горизонтальному переносу. Кислородному фотосинтезу предшествовал фотосинтез бескислородный, более простой, требующий участия одного белкового комплекса – «фотосистемы». Предки цианобактерий умудрились скомбинировать в своем геноме сразу две слегка различающиеся фотосистемы. Одна, вероятно, была у них «своя», а вторую они позаимствовали у другого бескислородного фотосинтетика. Комбинация двух фотосистем в одной клетке позволила цианобактериям перейти к кислородному фотосинтезу – процессу более сложному, но зато и более перспективному. Для бескислородного фотосинтеза требуются дефицитные вещества – доноры электрона, например сероводород или двухвалентное железо. При кислородном фотосинтезе донором электрона служит обычная вода – колоссальное облегчение!
Все это хорошо, но есть одна проблема. То, что мы видим в геномах прокариот, – все эти заимствованные у других микробов полезные гены – не результат ГПГ в чистом виде, а результат комбинации ГПГ и отбора. Мы видим только удачные переносы, потому что неудачные отсеяны отбором. Сколько их было? Точную цифру назвать трудно, но наверняка гораздо больше, чем удачных. Хватать без разбора чужие фрагменты ДНК и встраивать их в свой геном – занятие крайне рискованное. Шанс приобрести что-то полезное при этом ничтожно мал по сравнению с шансами повредить свой геном, испортить важный ген ненужной вставкой или заменой, приобрести что-то несовместимое с имеющимися у вас генами, заполучить смертоносный вирус или активный мобильный элемент, который начнет размножаться и прыгать как сумасшедший и превратит ваш геном в бессмысленную лапшу.
Полезный новый ген у неродственного микроба удается заполучить в среднем лишь раз-другой за миллион лет (примерно такие цифры дал анализ геномов бактерий и одноклеточных эукариот). При этом бесполезные и вредные новые гены доступны в неограниченном количестве. Их можно получать десятками ежедневно.
Естественный отбор не обладает даром предвидения. Он не будет «терпеть» вредный признак (склонность заимствовать чужие гены) миллион лет ради того, чтобы после всех перенесенных мучений приобрести наконец что-то полезное.
Из этого следует, что микробы сохраняют способность к заимствованию генов не потому, что надеются «одолжить» у неродственных микробов какое-нибудь замечательное новшество, новый полезный ген или генный комплекс. Отбор не смог бы поддерживать способность к ГПГ ради такого «журавля в небе». Остается одно из двух. Либо эта способность вообще не поддерживается отбором и является неким «неизбежным злом», либо микробы обмениваются генами по какой-то другой причине: более повседневной и насущной, связанной с какими-то сиюминутными выгодами. Первый вариант маловероятный, отбор умеет сводить «зло» к минимуму, а в природе почти все организмы практикуют генетический обмен. Значит, более осмысленным будет обсуждение второго варианта – выгоды здесь и сейчас. Что это за выгоды? Это мы сейчас и попытаемся выяснить.
Секс против вредных мутаций
Допустим, существует популяция микробов, не умеющих меняться генами (такие организмы называют бесполыми). Допустим, у каждого новорожденного микроба происходит одна вредная мутация. В этом случае популяция обречена на вырождение. С каждым поколением груз вредных мутаций будет расти, а приспособленность – падать. Никакой отбор не сможет остановить накопление мутационного груза, потому что все особи поколения N будут иметь по N вредных мутаций: отбирать некого. Единственное, что сможет сделать отбор, – это замедлить снижение приспособленности, отбраковывая более вредные мутации и сохраняя менее вредные.
Эту идею (в приложении не к микробам с их ГПГ, а к эукариотам с половым размножением – амфимиксисом) разработал великий эволюционный генетик Герман Мёллер (1890–1967). Она вошла в науку под названием «храповик Мёллера». Храповик – это устройство, в котором ось может крутиться только в одну сторону. Имеется в виду, что средняя приспособленность бесполой популяции под действием вредных мутаций может меняться только в сторону ухудшения. Например, если случайно погибнет или мутирует «лучшая» особь в популяции, то эта потеря необратима. Храповик повернулся на один щелчок. Ведь без секса невозможно собрать из пары «плохих» геномов геном получше.
Но если микробы умеют заимствовать чужие гены, то в каждом поколении найдутся счастливчики, которые заменят свой испорченный ген на его неиспорченную версию, взятую у другого микроба, у которого вредная мутация произошла в другом гене. Поэтому в каждом поколении, несмотря на мутагенез, какая-то часть особей окажется свободной от вредных мутаций. Их-то и поддержит отбор, они-то и оставят больше всего потомков. При достаточно мощном отборе популяция сумеет избежать вырождения.
Идею можно выразить иначе. Если у бесполого организма возникает вредная мутация, его потомки уже не смогут от нее избавиться. Она будет, как родовое проклятие, передаваться всем его потомкам вечно (если только не произойдет обратная мутация, что крайне маловероятно). У бесполых организмов отбор может отбраковывать только целые геномы, но не отдельные гены. Сойдут с арены те несчастные, которым совсем не повезло, а останутся те, кого мутационная судьба в этот раз пощадила (при этом вместе с вредной мутацией может исчезнуть и редкий полезный признак). Но и они окажутся хуже, чем их родители, просто их мутации не такие зловредные. Поэтому в череде поколений бесполых организмов вредные мутации могут неуклонно накапливаться. Храповик Мёллера – это настоящая трагедия отцов и детей: отцы могут справедливо сетовать на падение нравов.
Но если организмы размножаются половым путем (или хотя бы изредка практикуют ГПГ с гомологичной рекомбинацией), то индивидуальные геномы иногда перемешиваются. Новые геномы при этом собираются из фрагментов, ранее принадлежавших разным организмам. В результате возникает новая сущность, которой нет у бесполых организмов – генофонд популяции. Гены получают возможность размножаться и выбраковываться поодиночке, независимо друг от друга, а не в неразрывной связке с другими генами данного генома. Отбор получает возможность отделять зерна от плевел: ген с неудачной мутацией может быть отсеян отбором, а остальные гены данного родительского организма могут при этом сохраниться в генофонде.
Таким образом, секс помогает отбору очищать генофонд от постоянно возникающих вредных мутаций, тем самым спасая популяцию от вырождения.
Секс в защиту полезных мутаций
С полезными мутациями ситуация очень похожая. Допустим, в популяции бесполых микробов возникло две полезные мутации. У одного микроба произошла полезная мутация в гене А, у другого – в гене Б. Что будет с потомками удачливых мутантов?
Поскольку обе мутации полезны, потомки мутантов будут размножаться быстрее прочих особей (мы не рассматриваем усложненный вариант, когда оба полезных признака отягощены букетом других мутаций разной степени вредности). Каждый мутант даст начало быстро размножающемуся клону. В конце концов все немутанты будут вытеснены и в популяции останутся два успешных клона: один с мутацией в гене А, другой с мутацией в гене Б. Пока все идет не так уж плохо (с точки зрения «пользы» для популяции).
Дальше начинаются неприятности. Поскольку микробы бесполые, объединить обе мутации в одном геноме они не в силах. Вместо этого начнется конкуренция между двумя клонами, или, по-научному, клональная интерференция. Тот клон, чья мутация оказалась более полезной, в итоге победит, а второй клон (тот, чья мутация оказалась менее полезной) будет вытеснен, т. е. попросту исчезнет. Таким образом, из двух полезных мутаций зафиксируется только одна. Вторая будет утрачена, хотя ее очень жаль терять!
Неэффективность налицо. Бесполая популяция – весьма несовершенная «машина для эволюции». А все потому, что она даже и не популяция в полном смысле слова. Это свора изолированных клонов, жестоко конкурирующих друг с другом.
Но если мы предоставим нашим микробам возможность заимствовать чужие гены, то какой-то мутант с улучшенным геном А рано или поздно позаимствует у микроба из другого клона улучшенный ген Б (или наоборот). В результате появится микроб с обеими полезными мутациями вместе. Его-то потомки и унаследуют мир. Польза ГПГ очевидна, не правда ли? Поэтому если смотреть на проблему отцов и детей глазами эволюциониста, то она легко решается с помощью секса.
Секс делает все полезные мутации, возникшие в популяции, «общим достоянием». Поэтому скорость приспособления к меняющимся условиям у организмов, способных к сексу, должна быть выше, чем у бесполых. Эту идею первыми разработали в 1930-х годах уже упоминавшийся Герман Мёллер и другой гениальный генетик-эволюционист Рональд Фишер (1890–1962). Она так и называется – эффект Фишера – Мёллера.
Эффект Фишера – Мёллера тем сильнее, чем выше частота возникновения полезных мутаций. Эта частота, в свою очередь, зависит от условий среды. Чем хуже условия, тем чаще возникают полезные мутации, чем условия благоприятнее, тем это происходит реже. В этом нет никакой мистики, так получается автоматически. Ведь приспособленность организма и благоприятность условий – стороны одной медали. Ухудшение условий идентично снижению приспособленности. Чем ниже приспособленность (т. е. чем дальше находится организм от локального пика на ландшафте приспособленности), тем выше вероятность того, что случайная мутация окажется полезной. Из этого следует, что эффект Фишера – Мёллера должен быть сильнее в переменчивой среде, к которой организмы не успевают как следует приспособиться.
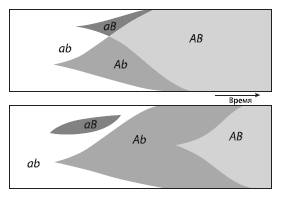
Схема, показывающая, как секс может ускорять распространение полезных мутаций. При половом размножении (верхний рисунок) два новых полезных аллеля (A и B) объединяются в результате скрещивания особей, каждая из которых имеет только один из этих аллелей. При бесполом размножении (нижний рисунок) приходится дожидаться, пока обе мутации случайно возникнут у одного и того же клона.
Рассмотренные модели применимы к любым популяциям, кроме чрезвычайно больших или абстрактных «бесконечно больших» популяций, с которыми любят играть специалисты по эволюционной генетике. Дело в том, что для бесконечно больших популяций удобно выводить красивые формулы. В популяциях с ограниченной численностью секс, по-разному комбинируя полезные и вредные мутации, регулярно создает генотипы с повышенной приспособленностью, появление которых в бесполой популяции маловероятно. Что касается бесконечно больших популяций, то там все не так очевидно: требуется соблюдение ряда дополнительных условий, чтобы секс давал ощутимое преимущество. Специалисты спорят, насколько часто соблюдаются эти условия в природе. Можно поспорить и о том, часто ли в природе встречаются настолько громадные популяции, что их можно считать бесконечными без ущерба для точности моделей. Общий вывод, впрочем, от этого не меняется. Как правило, секс полезен, особенно если за него не приходится слишком дорого платить.
Секс помогает извлечь пользу из численности
Из модели Фишера – Мёллера вытекает интересное следствие: польза от секса в большой популяции может быть больше, чем в маленькой. Выше мы рассмотрели случай, когда в двух популяциях – половой и бесполой – возникло по две полезные мутации. В бесполой популяции зафиксировалась только одна из них, а вторая пала жертвой безжалостной конкуренции между клонами. В популяции организмов, способных к сексу, зафиксировались обе мутации.
Задумаемся теперь, что произойдет, если мы вдвое увеличим численность обеих популяций.
Логично допустить, что в популяции с удвоенной численностью будет возникать вдвое больше редких полезных мутаций в единицу времени. Поэтому давайте предположим, что в каждой популяции возникло уже не по две, а по четыре полезных мутации (в четырех разных генах у четырех разных особей). Какая судьба их ждет? Ответ очевиден. В половой популяции все четыре мутации объединятся в одном геноме (мы предполагаем, что эффект мутаций аддитивен, т. е. их польза складывается и они не мешают друг другу). Положительное влияние численности налицо: вдвое больше численность – вдвое больше полезных мутаций зафиксировалось.
В бесполой популяции начнется конкуренция между четырьмя клонами – потомками удачливых мутантов. Победит тот клон, чья мутация окажется самой полезной. Остальные три полезные мутации будут вытеснены. Таким образом, почти никакой пользы от удвоения численности бесполая популяция не получит. Ну разве что самую минимальную – за счет того что на этот раз зафиксируется лучшая из четырех мутаций, а не из двух[39].
Итак, сексуальная популяция с единым генофондом будет адаптироваться тем эффективнее, чем выше ее численность. Бесполая «свора конкурирующих клонов» не получает такого выигрыша от увеличения численности.
В 2002 году этот теоретически предсказанный эффект удалось подтвердить экспериментально. Ник Коулгрейв из Эдинбургского университета работал с жгутиконосцами – хламидомонадами (Chlamydomonas reinhardtii). Эти одноклеточные водоросли могут размножаться как бесполым путем (делением), так и половым, образуя гаметы, которые затем сливаются в зиготы. Половое размножение у хламидомонад можно стимулировать искусственно (поместив жгутиконосцев в воду, не содержащую соединений азота), а можно, наоборот, заблокировать – например, поместив в аквариум жгутиконосцев только одного «пола» (вообще-то у них нет самцов и самок, зато есть так называемые типы спаривания, причем скрещивание возможно лишь между представителями разных «типов», которые ничем, кроме половой избирательности, друг от друга не отличаются). Коулгрейв заставил множество больших, средних и маленьких популяций хламидомонад приспосабливаться к неблагоприятным для них условиям. При этом половина популяций размножалась как бесполым, так и половым путем, а другая половина – только бесполым.
Спустя 50 поколений у всех подопытных популяций была измерена приспособленность (скорость размножения по сравнению с исходными, предковыми жгутиконосцами). Оказалось, что все бесполые популяции приспособились к новой среде почти одинаково плохо – большие лишь ненамного лучше маленьких. Популяции, практиковавшие секс, приспособились лучше, чем бесполые. Самое главное, чем выше была численность, тем сильнее проявилось их преимущество. Маленькие популяции (состоявшие примерно из 1000 особей) приспособились к новой среде лишь на 2 % лучше, чем бесполые, средние (100 тыс. особей) – на 7 %, большие (1 млн особей) – на 13 % (Colegrave, 2002).
Таким образом, проверяемое следствие, вытекающее из модели Фишера – Мёллера, замечательно подтвердилось. Разумеется, то, что справедливо для хламидомонад, не обязательно должно быть верным для всех живых существ. Но никто и не говорит, что в жизни все просто и однозначно.
Переменчивая среда способствует половому размножению
Еще одно подтверждение эффекта Фишера – Мёллера удалось получить в опытах на коловратках (Becks, Agrawal, 2010). Коловратки класса Monogononta (однояичниковые) способны как к бесполому (партеногенетическому), так и к половому размножению[40]. При низкой плотности популяции в ней, как правило, присутствуют только самки, производящие так называемые амиктические (партеногенетические) диплоидные яйца, из которых без оплодотворения выводится следующее поколение самок. При высокой плотности часть самок начинает производить гаплоидные яйца, из которых выходят маленькие непитающиеся самцы. Они спариваются с самками, в результате чего образуются «покоящиеся» оплодотворенные яйца с плотной оболочкой. Из них снова выводятся только самки. Решение о переходе к половому размножению коловратки принимают на основе так называемого чувства кворума. Это происходит, когда концентрация веществ, выделяемых самками, превышает определенный порог. Поэтому, чтобы спровоцировать самку на половое размножение, достаточно поместить ее в воду, взятую из аквариума, где плотность популяции коловраток высока.
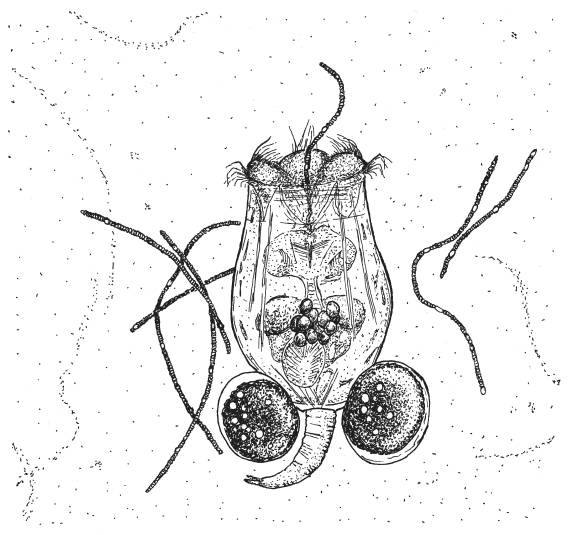
Коловратка Brachionus calyciflorus поедает колонию нитчатых цианобактерий Anabaena. Коловратка несет на себе два крупных партеногенетических яйца.
У коловраток Brachionus варьирует склонность к тому или другому способу размножения, и эти вариации наследственные. Это значит, что выбранный коловраткой способ размножения зависит не только от среды, но и от генов: встречаются клоны самок, легко и быстро переходящие к половому размножению, тогда как другие клоны делают это менее охотно.
Эти особенности делают коловраток удобным объектом для экспериментального изучения эволюции пола. Выше мы говорили, что эффект Фишера – Мёллера лучше проявляется в переменчивой среде: в этих условиях половое размножение становится более выгодным (Pylkov et al., 1998; Lenormand, Otto, 2000; Agrawal, 2009). Суть идеи в следующем. Допустим, популяция состоит из двух частей (субпопуляций), живущих в разных условиях, причем между частями существует обмен особями (миграция). В каждой из двух субпопуляций отбор благоприятствует разным комбинациям генов (точнее, генетических вариантов – аллелей). Если особь мигрирует из одной субпопуляции в другую, некоторые ее гены окажутся в новых условиях вредными, снижающими приспособленность. Поэтому всем остальным генам данной особи будет выгодно избавиться от этого груза и найти себе более подходящую «компанию», т. е. объединиться с генами аборигенов, лучше приспособленными к местным условиям. Этого легко добиться при помощи полового размножения, но совершенно невозможно, если вы размножаетесь партеногенезом. Поэтому, если миграции происходят достаточно регулярно, любая мутация, повышающая склонность особей к половому размножению, имеет шанс распространиться в популяции и вытеснить конкурирующий аллель, снижающий частоту полового размножения.