


Полная версия
Collins New Naturalist Library
The mesosoma is the motor part of the body and carries the six legs and, in sexuals, the two pairs of wings. Each leg has five main joints but the outermost one is finely subdivided to make a flexible foot that ends in two curved claws. The articulation and manoeuvrability of each leg is remarkable; they are perfectly constructed for movement over rough and irregular ground and up vertical and overhanging surfaces; they can be lifted and rubbed over the body for grooming or so that, in the case of the young queen, the wings can be broken off. The forelegs, besides having a comb, are used for holding food and are placed round the head of a larva when it is being given liquid. The hindlegs are the longest and can be used to lift the body well clear of the ground; in this way the gaster can be brought under the mesosoma and venom ejected forwards or, when the individual is laying an egg, it can be taken off with the mandibles.
The mesosoma of the queen is much bigger than that of the worker and is divided into plates joined by elastic sutures that play a part in controlling wing movement during flight. In workers nearly all the sutures are sealed over and the mesosoma is a single rigid structure. At the hind end there may be spines which are thought to protect the petiole from damage by predators and other aggressive ants and there are two large pockets that carry a grease which is said to be spread over the body, to be antiseptic and perhaps to carry a species-characteristic smell. The gland cells which produce this are just below the inner surface of the pocket. On each side of the mesosoma there are three spiracles. These are entrances to complicated systems of air tubes (tracheae) that ramify both forwards into the head and backwards into the gaster. Gases are carried in this way to and from the tissues which absorb oxygen and emit carbon dioxide.
The most striking external feature of the gaster is its extensibility (fig. 4). Supple zones of cuticle are folded and tucked under tougher, but still comparatively flexible, zones in such a way that each segment telescopes into the one in front. A fully distended gaster shows the intersegmental cuticle as white bands between much darker segmental ones. Right at the tip is the anus, often surrounded by a ring of hairs and below this there is an opening for eggs. The only appendage is the sting and its sheath but in many groups of ant even these are missing. The sting is a simple, pointed object without barbs (such as occur in the honeybee). It is tough and quite flexible, not brittle and can be gradually pushed into the soft body parts of insects and through the skin of birds and mammals, provided the ant can get a firm grip. Queens have stings like workers but very little is known about how and when they use them. They do fight each other under certain conditions and may use their venom like honeybee queens do; it is most likely, however, that stings and venom are used in defence of brood during the early stages of colony foundation when there are too few workers to defend the nest unaided.
At the front of the gaster (in myrmicine ants) there is a sound-producing organ (fig. 4). The second petiolar segment has a very stiff hind margin that can be rubbed against a series of fine ridges in the cuticle on the dorsal surface of the gaster. When the gaster is lifted up and down a squeak is produced. It is just audible in ants of the genus Myrmica if they are held firmly by the head and thorax leaving the gaster free to move. Curiously, this sound seems to have no social function at all; other ants ignore it. The only plausible suggestion is that it shocks predators that have acute hearing and causes them to drop the ant, which then has a chance to escape.
In the integument, particularly at joints, there are many microscopic sense organs that send into the central nervous system information about the stance and position of the body relative to the substratum. These enable the ant to assess its relation to gravity and provide it with information important in navigation.
INTERNAL FEATURES
Gases are taken to and from the tissues in air tubes from the six spiracles which have already been mentioned. Fluids bathe all the internal organs and are circulated by a long, thin, tubular pump (the heart) which runs dorsally all the way from the tip of the gaster to the inside of the head just behind the brain; it has valves along the way which prevent back flow (fig. 4). These fluids then filter through the organs and tissues. Blood does not carry gases and is not piped to and from the tissues, as is the case in vertebrates.
Much of the head is occupied by muscles which work the jaws and the food pump (pharynx). The biggest muscles are those that close the mandibles and they are obviously capable of exercising a strong and tight grip; the muscles are fixed on the back of the head and run to the inner edge of each jaw (fig. 3). Seated on the ventral part of the head and running to the outer part of each jaw are the muscles which open them; they are much smaller. Other small muscles retract the tongue after it has been extended by blood pressure and there are some inside which manipulate it. The pharynx is a flattened part of the gut in the front of the head which can be enlarged and diminished by means of six muscles attached to it in various ways, some from above, some from the sides and some transversely (fig. 2). It pumps liquid food up after the silt and solid particles have been taken out and passes it down the oesophagus. This latter is a very thin tube, more or less circular in cross section that runs right through the mesosoma and petiole to the gaster. The brain is a ganglion of nerve cells and fibres behind the pharynx and over the oesophagus. It collects sensory data from outside and inside the body, works out correlations and associations and stores relevant information. This enables the insect to learn and react to quite complicated patterns of stimuli. The brain also has closely associated with it a number of ductless glands that control most internal chemical processes. Underneath the oesophagus is another ganglion, connected to the brain by thick nerve trunks, which is largely responsible for controlling motor activity; it co-ordinates the movements of antennae and mouth parts. In its turn it sends nerve trunks into the mesosoma.
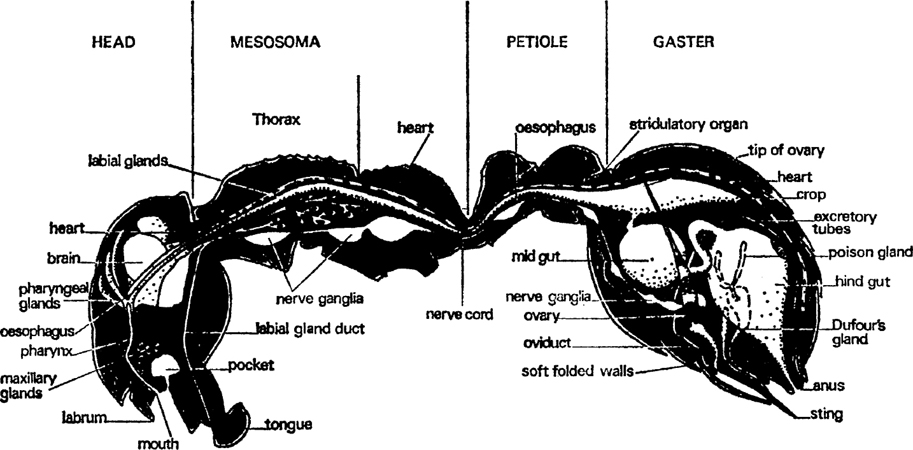
FIG. 1. Worker of Myrmica rubra, sagittal section showing main organs and divisions of the body (after Janet).
A gland opens on each mouth part (figs. 1, 2, 3). The mandibles have glands that lie against the outer wall of the head, one on each side between the eye and the mandible base. A single layer of cells produces a secretion which is stored in a thin-walled reservoir and conducted by tube to the base of the mandible where it opens through a pore. These glands have several functions: they are partly digestive, partly they lubricate the joints of the mouth and they also contain volatile substances that alert other ants. The maxillary glands have no reservoir and open directly at the base of the maxillae; they probably produce a digestive fluid. The labial glands are not in the head at all; they occur in the front of the mesosoma and are connected to the tip of the tongue by a long, thin tube. They supply a watery lubricant which contains some digestive juices. The most striking gland in the head, next to the mandibular, is the pharyngeal gland; this consists of about twenty finger-shaped tubes, often containing a yellow oil. It is thought that this substance is separated from the food as it travels through the pharynx; it may be digested and used as an energy supply. The watery part of the food goes down the oesophagus to the crop which is in the gaster. From here it can, nevertheless, be regurgitated to larvae and queens and as it contains most of the proteins it is of great value to them, either for body growth or egg production, as the case may be.
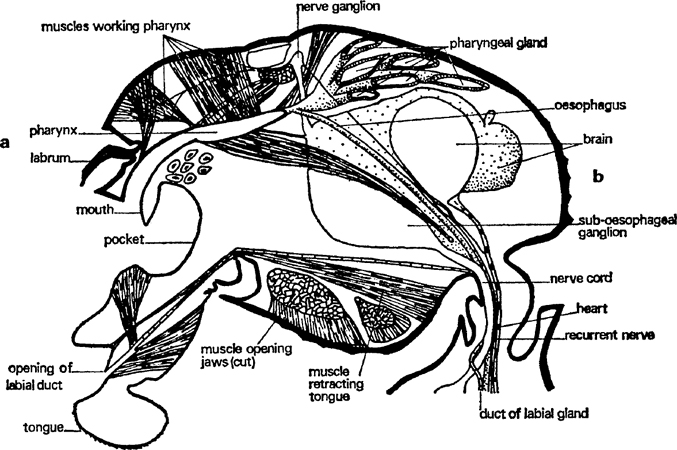
FIG. 2. Worker of Myrmica rubra, head sagittal section showing main organs (after Janet).
In the narrow constriction between the head and the mesosoma there are six channels: the ventral nerve cord, the duct of the labial gland, the oesophagus, two air ducts and the tubular heart. In the mesosoma the ventral nerve cord forms into three ganglia (the last a compound one) that are largely concerned with co-ordinating leg and wing movements. It contains all the muscles operating the legs and, in the sexuals, the wings. In fertile queens these wing muscles degenerate and the material from them is used to make eggs which not only give rise to a brood of larvae but are also used to feed them. Later on, when the colony is well established, the space occupied by the muscles becomes filled with storage tissue called fat-body from the opalescent oil droplets that are the principal component. Muscular action uses a lot of energy and the mesosomal organ must be well-supplied with oxygen from the three pairs of air tubes. There is also a pair of large air sacs which may be ventilated during muscular movement in some way.
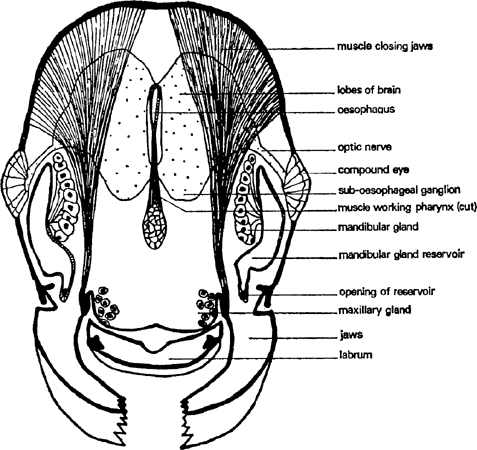
FIG. 3. Worker of Myrmica rubra, head section from a. to b. of fig. 2 (after Janet).
Water-dissolved substances are carried through the long, thin oesophagus to be stored in a dilatation called the crop which lies dorsally in the front of the gaster. This can be distended to a huge size for it is used to carry nectar and honeydew back to the nest. There is a valve between the crop and the rest of the gut which prevents a good deal of loss from the crop but not all. In some ants this is held closed by muscular sphincters; in others it closes passively when the fluid pressure in the crop rises. It is thought that this second method of control is more efficient since it uses no energy. The crop contents are mostly very liquid and can be regurgitated to other adults and to larvae. This is probably done by telescoping the gastral segments inwards but perhaps by reversing the pharyngeal pump mechanism.
Once fluid has passed back into the midgut it can no longer be shared by other ants. The thick walls of the midgut (fig. 4) secrete digestive enzymes and absorb the small molecules into which the food is broken. Waste material passes back through another tube into a thin-walled hindgut which stores indigestible residues and probably extracts some water and water-soluble substances. Finally, the liquid waste is ejected in a special part of the nest some way away from the brood rearing zone, along with the refuse. Foragers probably eject their own residues outside the nest. It includes waste substances extracted from the body fluids by long, thin, hollow, thread-like tubules; they join the gut just after the midgut. Much of this excretion is synthesized into uric acid which appears as white granules. In workers the main storage tissue is in the gaster. Oils, glycogen and proteins are all stored there though most of it is oil.
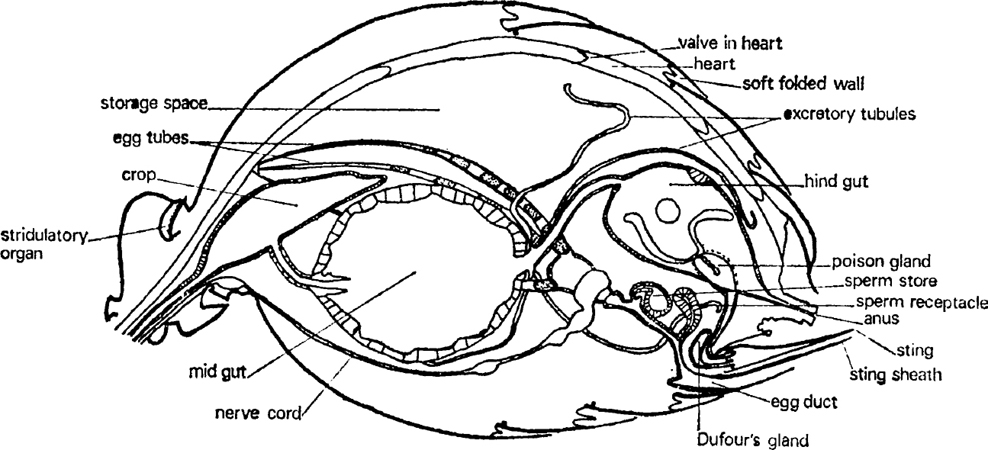
FIG. 4. Queen of Myrmica rubra, gaster sagittal section showing main organs (after Janet).
A very important part of the gaster is occupied by the genital system (fig. 4). This consists of thin-walled egg tubes (many more in queens than in workers) that start just below the heart at the front of the gaster and pass backwards and downwards to the genital opening below the anus and the sting. Eggs start as small cells at the tip and move down, growing and maturing as they go. Part of the material for growth comes from nurse cells which are broken down and incorporated into the egg tissue but some is extracted from the blood by an enveloping layer of follicle cells. When the egg has been fully formed it is enclosed in a special skin (the chorion) and the egg passes into the oviduct to be fertilized. The oviduct is a simple tube with a thicker wall than the ovary. In queens it has two pouches on its dorsal side, one near the outside which received sperm during copulation and the other a bit farther in that stores the sperm alive for a decade or more. How it gets from one to the other is unknown. These pouches are not present in workers unless the ant is a very primitive species.
It remains only to mention two glands that open into the sting (fig. 4). One is the poison gland which consists of two closed tubes feeding a thin-walled storage vesicle and which contains a watery solution of mixed venoms. The other is a smaller gland called after Dufour which contains an oily material secreted by cells which surround the storage space. The poison glands vary quite considerably in different families of ant, depending upon whether they synthesize a thin liquid for squirting or injecting with a sting, or a sticky fluid for smearing.
In general the ant body is neatly divided for separate functions: the head carries the main sense organs and the brain and has in front a mechanism for food capture and food pre-treatment, the mesosoma is specialized for locomotion and the gaster has a region of absorption and food storage but also contains the reproductive organs. Defensive apparatus is disposed both in front and behind but the body is flexible enough for both to act forwards in concert.
LARVAL STRUCTURE
Ants’ eggs are oval and white; each egg weighs only ·00005 gm and is less than a millimetre long. In most of them a legless grub develops. This is of course no bigger than an egg at first and breaks its way out by means of its sharply pointed jaws. It is almost hairless and transparent so that a small, residual blob of egg yolk in its midgut can easily be seen. Though its cuticle is inelastic rather like polythene it is wrinkled and folded so that a good deal of room for expansion is allowed. There are three stages separated by moults, when the old skin is cast off and a new one, folded and wrinkled to allow for growth, is built underneath. Only the head has a firm cuticle that will not expand at all so that its growth only takes place during the moult between successive stages. The second stage has a good many more hairs than the first and the third and last stage is very hairy indeed. It has some hooked hairs as well as the simple ones which enable the grubs to interlock and so be carried about in one group. It is tempting to believe that the workers are able to distinguish the stages by feeling the degree of hairiness with their antennae, but again this has not been tested by experiment.
The head carries jaws with pointed, hardened tips. These are capable of piercing eggs and can masticate soft tissues a little. There are several sense organs of a very simple kind, including minute, unjointed, conical antennae; several other similar ones occur around the mouth and are probably used by the larva to feel and taste what it is eating, as well as to enable it to lock on to the worker’s mouth whilst it is being fed regurgitated liquid. The head, like that of the adult, contains a pharynx innervated by muscles (fig. 5). It is a pump that only sucks inwards; larvae cannot regurgitate food. The ‘brain’ consists of a dorsal ganglion and a ventral ganglion with nerve trunks running round the oesophagus to connect them. The ventral ganglion is the first of a long, chain-like series that runs down the body near the ventral wall.
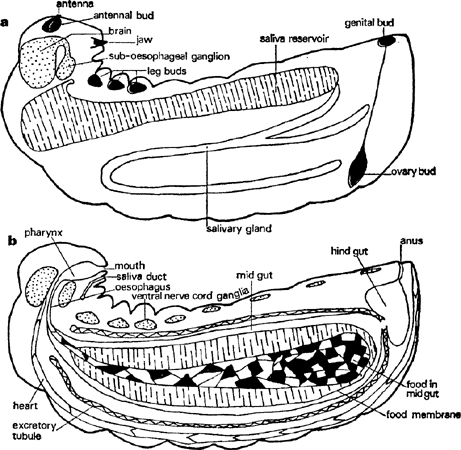
FIG. 5. Larva of Myrmica rubra in third instar, longitudinal sections to show main organs: a. slightly off mid-line, b. on mid-line (sagittal).
Most of the body contents are connected with the digestion and storage of food. A very large, sac-shaped midgut consists of a single layer of large, glandular cells. This is connected to the pharynx by a thin-walled oesophagus and where this and the midgut meet there is a structure for secreting and moulding a thin membrane which encloses the food and is thought to protect the delicate gut cells from abrasion. However, there is no way through from the midgut into the much smaller hindgut and all the food residues collect and condense into a black pellet surrounded by food membranes which are ejected at the end of the larval stage. The hindgut is very thin-walled and collects the liquids extracted by excretory tubules that lie freely in the body fluids. It thus acts as a bladder by storing urine and, if the diet is rich in protein, white insoluble granules of uric acid. The bladder is emptied by a longitudinal contraction of the body; this usually occurs only in response to a light touch on the skin in the region of the anus so that normally there is a worker nearby to collect it up and carry it away. The larva has a single but large pair of tubular digestive glands that start in the front, pass backwards and then enlarge into thin-walled reservoirs which go forwards again well into the first body segment just behind the head before sending tubes that join and run to the lower lip. A great deal of fluid that is rich in protein-splitting enzymes is stored in these reservoirs, ready to be shed on food: as they are thin-walled and quite close to the body wall they are really a considerable hazard for if they are broken in an attack the juices escape into the larval body and digest its internal organs. Saliva may be ejected as a droplet if the larva’s head is pushed backwards.
A long, thin, tubular, valved heart runs mid-dorsally from just above the anus to just behind the brain and circulates body fluid, as in the adult (fig. 5). There are a number of muscles inside each larva which enable it to make some simple movements, such as bringing its head down towards its abdomen or retracting it into the thorax, moving its jaws and ejecting urine but they also contract in a way that maintains blood pressure against the body wall and hence turgidity. A large part of the body space is filled with storage tissue, called fat-body from the abundance of oil in it; both glycogen and protein are also stored there.
Rudiments of the adult are distributed throughout the larva in appropriate positions (fig. 5). At first they are simple, hollow sacs surrounded by thin-walled sheaths but as they grow they elongate and split transversely or longitudinally into segments. Thus the antennae are situated in front of the brain and are nearly spherical. The six leg buds are very similar in shape and are arranged in three pairs on either side of the nerve cord in the first three body segments. All of these grow in length and split transversely. The wings are buds, again almost spherical at first, which lie on the side of the second and third segments. They retain a flattened form but develop into a heart-shaped one by the growth of their ventral tips. The ovaries are exceptional. They are paired, solid, carrot-shaped organs fixed by the broad end to the heart not far from the end of the abdomen; their other end passes as a thin filament right down to the genital buds just in front of the anus.
When the chemical signal for metamorphosis is given these buds grow more quickly and begin to join up and assume their adult shape. Even the sheaths spread out and form, at least in the legs and antennae, some of the basal segments. The wings elongate but never split. The ovary, in the case of a queen-forming larva, splits longitudinally into about eight parts which grow along the filament towards the genital buds where they meet the oviduct as it grows upwards. In the worker this is a narrow tube and a broader one in the queen.
All this takes place in the larval skin after the larva has stopped feeding and ejected its food residues through the hindgut which breaks down where it touches the midgut. The membrane enclosing all the residues not only makes this easier but probably prevents gut bacteria getting into the body and starting an infection. The larval head is not big enough to hold the new pupal head and so this is formed in the first body segment; only the tips of the pupal antennae lie in the larval head. The young pupa escapes through a split in the larval skin which starts dorsally just behind the head. Somehow or other it manages to push off the larval skin as far as the tip of its gaster without any help from workers. Simultaneously the petiole is formed by a contraction of the second and third abdominal segments. This together with the inward telescoping of the last body segments seems to reduce the body volume and create enough blood pressure to inflate and smooth out the surface just before it sets.
Only one more moult is needed before the adult is produced. This forms with much less change in shape inside the pupal skin. The wings flatten, elongate and fold up and the legs and antennae and petiole segments narrow a little more. Colour appears as a gradually increasing brownness in the general body but the eyes change from pink to black. The adult emerges with a very soft flexible skin and the wings, if any, inflate, spread out and set hard.
The caste differences in the female do not develop until very near the end of larval life. In workers the wing buds stop growing when they are quite small and all traces vanish by the time the pupa is formed; their ovary never thickens and splits into egg tubes but simply elongates to meet the oviduct. Caste differences in ants thus depend quite simply on whether buds grow or not; there is no degeneration and reorganization as happens in the honeybee. It seems likely that the growth of wings and ovaries is specially delayed during the development of females so that caste determination can be left to the last moment.
In males, which have only one caste, the wing grows earlier and the testis is split longitudinally very early in the larval stage. Late caste determination presumably gives a more sensitive response to social conditions.
Males are either the same size or smaller than the females and they are usually darker. Their antennae are straighter and clumsier and cannot be folded back against the head. They have well-developed compound eyes and ocelli. Their thorax, like that of the sexual females, carries two pairs of wings linked by hooks and it is composed of plates separated by sutures but their petiole is not quite so well-developed. There is no sting at the end of the abdomen; in its place are appendages for locking on to the female during copulation. They do not possess large body reserves like the sexual females.
CHAPTER 3
TYPES OF BRITISH ANT
IDENTIFICATION
Three keys are given here: the first to sub-families, the second to genera and the third to species. These are based on characteristics shown by workers and, in those species which lack them, queens; males help a great deal but for simplicity have been left out. To identify ants as far as the species is difficult; indeed, experts are often not in agreement about some very similar forms. Nevertheless the four main genera, Myrmica, Leptothorax, Lasius and Formica, have been here divided into their most common and easily identified species. The key is not based solely on structural features but includes in a few places reference to habitat, nest site and shape and other aspects of natural history. A low-powered stereoscopic microscope is best for assessing many characteristics but a hand lens may sometimes be adequate. Many distinctions are comparative, e.g., hairy or not hairy, and it is obvious that in these cases experience and reference to a reliable, modern collection is essential.
Key to sub-families
1Waist of 2 small segments (fig. 1)Myrmicinae—Waist of 1 segment22Gaster constricted between segments 1 and 2, with a well-developed stingPonerinae—No constriction, no sting33Five segments of the gaster visible from above, a circular orifice for ejecting venom fringed by guide hairs (fig. 7)Formicinae—Four segments of gaster visible from above, no circular orifice but a slit through which viscous defensive fluid is passed, no hairsDolichoderinaeKey to genera
