


Полная версия
Психофизиология. Психологическая физиология с основами физиологической психологии. Учебник
Уже отмечалась важная роль мембраны в передаче информации в мозге. Мембрана представляет собой барьер для прохождения нервного импульса. Именно поэтому связи между нейронами опосредуются химическими передатчиками – нейромедиаторами (mediator – посредник, англ.), выделяющимися из окончаний аксонов в области специализированных межклеточных контактов – синапсов. Синапс представляет собой мембраны двух соседних нейронов (передающего информацию и получающего ее) и пространство между ними, которое называется синаптической щелью. Синаптическая щель – это пространство шириною около 20 нм между мембранами пресинаптической (мембрана нейрона, находящегося перед синаптической щелью) и постсинаптической (мембрана клетки, находящейся после синаптической щели) клеток (рис. 1.15).
Различают аксо-соматические синапсы, сформированные мембранами аксона и тела (сомы) нейрона, аксо-дендритные, состоящие из мембраны аксона и дендрита, и аксо-аксональные, при которых аксон подходит к другому аксону. Синапс между аксоном и мышечным волокном называется нейромышечной кольцевой пластинкой.
Молекулы медиатора находятся в везикулах – особых пузырьках, расположенных в аксональной терминали (окончании аксона). ПД, достигая аксональной терминали, становится сигналом открытия кальциевых каналов, которые вызывают синхронный эндоцитоз – координированное выделение медиатора из везикул и поступление их в синаптическую щель. Медиатор связывается с рецептором, находящимся на постсинаптической мембране, который инициирует в постсинаптической клетке те или иные изменения в зависимости от вида рецептора. Медиатор, взаимодействуя с рецептором, может способствовать открытию ионных каналов (натрий-калиевых или кальциевых) или через аденилатциклазный механизм активировать внутриклеточного посредника – цАМФ (циклический аденозинмонофосфат) и цГМФ (циклический гуанозинмонофосфат). При открытии натрий-калиевого канала натрий поступает внутрь клетки, что приводит к деполяризации участка мембраны постсинаптического нейрона. Каждый синапс делает лишь незначительный вклад в этот процесс. Однако каждый нейрон непрерывно интегрирует до 1000 синаптических входов, которые суммируются нелинейно (рис. 1.16) и при достижении порогового потенциала вызывают ПД, т. е. распространяющийся вдоль аксона потенциал.

Рис. 1.15. Структура синапса (а); фотография синапса, увеличенного под электронным микроскопом в тысячи раз (выполненная D. D. Kinkel) (б); фотография аксональных терминалей на теле аксона (в) (Kalat, 1992).

Рис. 1.16. Временная и пространственная суммация импульсов (Kalat, 1992).
Синаптическая задержка – время между началом пресинаптической деполяризации и постсинаптической реакцией – составляет 0,5 мс. Всего лишь 1/10 часть этого времени обусловлена диффузией (проникновением) медиатора через синаптическую щель к постсинаптической мембране. Большая же часть времени тратится на открывание Са2+-каналов, через которые кальций проникает в аксонную терминаль и способствует высвобождению медиатора из везикул. Кальций в аксонной терминали имеется в крайне малых количествах. После того как он окажет свое действие, он удаляется, либо связываясь со специальным белком – кальмодулином, либо проникая в эндоплазматический ретикулум. Освобожденный медиатор может действовать как на ауторецепторы (расположенные на пресинаптической мембране), так и на постсинаптические рецепторы.
Согласно принципу английского ученого Г. Дейла, постулирующего метаболическое единство нейрона, во всех окончаниях нейрона выделяется один и тот же медиатор. В настоящее время доказано, что этот принцип касается только пресинаптического единства нейрона. Эффекты, которые вызываются данным медиатором, могут быть различны и зависят от клеток-мишеней (в данном случае постсинаптических нейронов). Знак синаптического действия – повышение постсинаптического потенциала или его падение – определяется не медиатором, а свойствами рецепторов на постсинаптической клетке.
Постсинаптические рецепторы одного пресинаптического нейрона могут фармакологически различаться и контролировать разные ионные каналы. Одна постсинаптическая клетка может иметь более одного типа рецепторов для данного медиатора, и каждый из этих рецепторов контролирует отличный от других механизм ионной проводимости.
Кроме нейронов, суммирующих и передающих информацию к другим клеткам, описаны так называемые пейсмекерные нейроны, способные самостоятельно генерировать электрические импульсы (Alving, 1968). Активность таких нейронов характеризуется синусоидальными колебаниями частотой 0,1–10 Гц и амплитудой 5–10 мВ. Эти нейроны при отсутствии любого внешнего воздействия обеспечивают периодическую генерацию ПД и передачу возбуждения другим нейронам.
Медиаторы
В начале XX века группа английских физиологов, возглавляемая Дж. Лэнгли, показала, что электрическая стимуляция вегетативных нервов вызывает изменения в органах, иннервируемых этими нервами. Оказалось также, что такие изменения можно вызвать инъекцией в организм экстрактов надпочечников. Дж. Лэнгли предположил, что клетки, иннервируемые вегетативными нервами, имеют две рецептивные субстанции – тормозную и возбуждающую.
На основании этих данных Т. Эллиот в 1905 г. выдвинул предположение, что возбуждающие импульсы в вегетативных нервах вызывают выделение адреналина. В 1921 г. австрийский ученый О. Леви обнаружил, что тормозное влияние блуждающего нерва на деятельность сердца опосредуется специфическим веществом, позднее идентифицированным как ацетилхолин. Г. Дейл привел веские аргументы в пользу того, что ацетилхолин является медиатором в вегетативных ганглиях и нервно-мышечных соединениях. Однако доказать наличие синаптической передачи с помощью медиатора, а не электрического потенциала стало возможным только в 50-х годах, когда исследователи начали использовать микроэлектроды и электронный микроскоп.
Все медиаторные соединения – это низкомолекулярные водорастворимые (дипольные) амины или аминокислоты и родственные им вещества. Ацетилхолин и катехоламины синтезируются из циркулирующих в крови предшественников, тогда как аминокислоты и пептиды в конечном счете образуются из глюкозы. Свидетельством консерватизма живой природы является то, что, несмотря на различие циркуляторных систем и метаболических путей, беспозвоночные и позвоночные животные в равной степени используют большинство общих медиаторов (табл. 1.3).
Таблица 1.3.
Характеристика некоторых медиаторов

Число пептидов, для которых доказаны медиаторные свойства, постоянно растет. Многие из этих веществ содержат от 2 до 10 аминокислот, что соответствует размеру, с одной стороны, мелких аминокислотных медиаторов, с другой – гормонов. Обилие пептидов создает впечатление неоднородности этой группы веществ. В то же время нарастающая информация о их роли в организме позволяет увидеть универсальные принципы их действия. Предполагается, что нейроэндокринные клетки, секретирующие пептиды, первыми появились в эволюции примитивных нервных систем. По-видимому, нейропептиды, производимые ими, достаточно консервативны, поскольку, как уже упоминалось, одинаковые вещества или близкие последовательности аминокислот обнаружены у филогенетически различных ветвей животных – беспозвоночных и позвоночных. Многие из них найдены не только в мозге, но и, например, в кишечнике. Есть предположение, что все пептид эргические клетки связаны общностью эмбрионального происхождения. Пептиды по сравнению с другими медиаторами оказывают свое действие в чрезвычайно низких концентрациях.
С тех пор как в 1921 г. был идентифицирован первый медиатор, число их в арсенале науки постоянно увеличивается и в настоящее время составляет около 50. Многие биологически активные вещества имеют сходную с ними структуру. Они могут усиливать действие медиаторов (такие вещества называются агонистами) или подавлять их активность (антагонисты). Например, лекарственные препараты, снимающие тревогу (седуксен и др.), усиливают действие тормозного нейромедиатора – гамма-аминомасляной кислоты. Антидепрессанты (например, прозак) являются агонистами серотонина. Кокаин усиливает действие дофамина. Он связывается с белком, удаляющим дофамин из места его активности, тем самым увеличивая время его действия. Никотин активирует рецепторы ацетилхолина. Энкефалины и эндорфины являются природными лигандами морфиновых рецепторов: в норме именно они связываются с рецепторами, с которыми в особых условиях – при употреблении наркотика – взаимодействует морфин.
Важным звеном в нервном пути, который служит мишенью всех наркотиков – амфетамина, никотина, алкоголя и опиатов, – является небольшая часть базальных ганглиев, называемая nucleus accumbens (прилежащее ядро). Антипсихотические препараты (нейролептики) предотвращают связывание дофамина с его рецепторами. Содержащие дофамин нейроны, находящиеся в области вентральной покрышки среднего мозга, посылают свои аксоны в префронтальную кору и базальные ганглии, которые участвуют в двигательном контроле. Это обусловливает побочное действие длительно применяющихся нейролептиков, связанное с развитием дискинезий.
В настоящее время описан еще один класс посредников, имеющих чрезвычайно малые размеры молекул. К ним относят оксид азота (NO) и оксид углерода, или угарный газ (CO). Оксид азота опосредует действие ацетилхолина при расширении сосудов, в том числе сердечной мышцы. Именно он является активным компонентом нитроглицерина, используемого для расслабления резко суженных венечных сосудов сердца при стенокардии (грудной жабе). Этот посредник обнаружен в 2 % клеток мозга. Он крайне токсичен, поэтому используется макрофагами (одним из видов клеток иммунной системы) для уничтожения бактерий, проникающих в организм. Второй посредник – угарный газ – не менее токсичен и опасен для человека. В мозге он выполняет важную роль, активируя вторичный (клеточный) посредник – цГМФ.
Белки, служащие рецепторами нейромедиаторов, можно разделить на два класса в зависимости от механизма их действия. К одному классу относятся белки ионных каналов, меняющие свою форму и открывающие каналы, по которым проходят ионы. Рецепторы другого класса располагаются по соседству с мембранными G-белками, разрывающими богатую энергией фосфатную связь в молекуле гуанозинтрифосфата, что инициирует каскад биохимических процессов, ведущих к специфической клеточной реакции через вторичные (клеточные) посредники. Эффекты, производимые этими белками, характеризуются медленным началом действия и большей продолжительностью, по сравнению с реакциями, связанными с открытием ионных каналов.
Функции нейрона
В настоящее время можно говорить о наличии трех основных функций нейрона. Наиболее распространенной является суммация возбуждающих и тормозных синаптических потенциалов и передача возбуждения следующему нейрону.
Описаны нейроны (прежде всего нейроны гипоталамуса), обладающие секреторной функцией. Они синтезируют биологически активные вещества – статины и либерины – и выделяют их в кровеносные сосуды воротной системы гипоталамуса. С током крови эти вещества попадают в переднюю долю гипофиза и способствуют реализации или накоплению его гормонов.
Наконец, существуют нейроны, обладающие спонтанной ауторитмической активностью. Их называют пейсмекерами, или водителями ритма. Эндогенные процессы подобных нейронов приводят к периодическому изменению ионной проницаемости мембраны и генерированию ПД. Взаимодействуя с другими клетками, они синхронизируют активность этих клеток.
Типы нервных волокон
По скорости проведения импульса и строению нервные волокна разделяют на три группы – A, B и C. Волокна типа А делятся на 4 подгруппы: альфа-, бета-, гамма-, дельта-. Альфа-волокна имеют самый большой диаметр (12–22 мк) и обладают наибольшей скоростью проведения возбуждения (70–120 м/сек). Такие волокна проводят информацию к скелетным мышцам и от них в мозг, что позволяет человеку достаточно быстро приспосабливать положение своего тела к ситуации. Остальные нервные волокна имеют меньший диаметр (2–12 мк) и соответственно меньшую скорость проведения импульса. Они несут информацию от сенсорных органов. Гамма-волокна передают возбуждение от моторных нейронов спинного мозга к интрафузальным мышечным волокнам.
Таблица 1.4.
Свойства различных нервных волокон теплокровных (Бабский, 1972).

К волокнам типа В относятся миелинизированные волокна, преимущественно преганглионарные, распространенные в автономной нервной системе. Скорость проведения информации по ним составляет 3–14 м/сек.
Волокна типа С – это немиелинизированные волокна, скорость проведения информации в них составляет 2–6 м/сек. Большинство из них являются постганглионарными волокнами автономной нервной системы. Эти волокна проводят также сигналы о боли, тепле, давлении (табл. 1.4) (Бабский, 1972).
Словарь
Вегетативная нервная система —
иннервирует гладкую мускулатуру и управляет деятельностью внутренних органов, поэтому ее также называют висцеральной.
Везикулы —
особые пузырьки, расположенные в аксонной терминали, в которых хранится медиатор.
Гематоэнцефалический барьер —
уникальный защитный слой между нейроном и кровеносным сосудом, образованный глиальными клетками.
Глия —
функционально разнообразные клетки мозга, способные делиться, но не участвующие в процессе передачи информации.
Модуль —
объединение нейронов коры, несущих одну функцию, в виде колонок. Модуль может включать более 100 тыс. клеток, огромное большинство которых образует локальные нейронные сети, выполняющие ту или иную функцию.
Нейрон —
основная функциональная клетка мозга, участвующая в передаче и хранении информации. Это поляризованная клетка, которая с помощью сильно разветвленных многочисленных отростков – дендритов – получает сигналы и через длинный неразветвленный отросток аксон посылает информацию другой клетке.
Периферическая нервная система —
состоит из соматической и вегетативной (автономной).
Соматическая нервная система —
обеспечивает контроль сокращений поперечно-полосатых мышц, т. е. всей скелетной мускулатуры. Ее нейроны находятся в передних рогах спинного мозга, а их аксоны через передние корешки спинного мозга иннервируют скелетные мышцы.
Потенциал действия —
временное изменение мембранного потенциала нейрона под действием приходящих сигналов. Распространяется в обоих направлениях от места возникновения (аксонного холмика).
Синапс —
специализированный межклеточный контакт. Представляет собой мембраны двух соседних нейронов (передающего информацию и получающего ее) и пространство между ними, которое называется синаптической щелью.
Спинной мозг —
можно рассматривать как продолжение заднего мозга; является центральным коммутатором (переключателем), передающим сообщения из ЦНС на периферию и обратно.
Центральная нервная система —
включает структуры, расположенные внутри черепа и позвоночника: головной и спинной мозг. Все, что находится вне этих костных структур, относится к периферической нервной системе.
Контрольные вопросы
1. Что такое психофизиология?
2. В чем единство и различие психофизиологии и физиологической психологии?
3. Структуры центральной нервной системы.
4. Периферическая нервная система. Симпатическая и парасимпатическая системы.
5. Нейрон и его функция.
6. Типы глиальных клеток.
7. Передача информации в ЦНС.
8. Синапс и синаптическая передача.
9. Что такое медиатор?
10. Характеристика наиболее известных медиаторов.
11. Типы нервных волокон.
Глава 2
Методы получения психофизиологической информации

Н. Н. Герардов. Плакат театрализованного бала. Музей 10. Художественные собрания СССР: сборник статей. Искусство русского модерна. Сост. А. С. Логинова М.: Советский художник, 1989
Р. У. Эшби (1964) сформулировал положение о том, что сложность объекта исследования предопределяет сложность метода его исследования. Это положение получило название принципа адекватности. Согласно ему, неправомерно изучать сложный объект, каковым является нейрофизиологический механизм психической деятельности, элементарными методами.
Регистрация психофизиологических параметров
По-видимому, самым адекватным прибором для оценки психофизиологических реакций будет другой человек, органы которого в процессе эволюции приспособились для этого. Общаясь, люди ежедневно пользуются этим «прибором», чтобы по поведенческим признакам определить мысли и чувства другого. В то же время каждый человек вносит в свою оценку «шум», зависящий от его воспитания, особенностей развития, наличия или отсутствия болезненных отклонений и так далее. То, что величина такого «шума» у людей различна, мешает использовать этот природный инструмент в качестве экспериментального (рис. 2.1).
Созданные человеком приборы также вносят «шум», но он зависит от конкретных причин, которые можно определить и вычесть из результата, что и делает их более эффективными в исследовании.
Многие физиологические процессы являются по своей сути электрохимическими, поэтому их можно зафиксировать, приложив электроды непосредственно на изучаемый участок тела. Первые доказательства электрической природы физиологических процессов обнаружил Луиджи Гальвани в 1786 г., назвавший эту движущую силу «животным электричеством». Он искренне верил в особые качества этого электричества по сравнению с электричеством, открытым до него физиками (Хэссет, 1981). Дальнейшие исследования не подтвердили его предположения.
Обычно величина регистрируемых физиологических реакций в норме очень невелика, поэтому их усиливают и затем передают на записывающее устройство. Те физиологические процессы, которые имеют отличную от электрической природу, предварительно преобразуют в электрический сигнал, чтобы затем зафиксировать.

Рис. 2.1. Э. Руссо. Ритуалы и мифы нашего времени (обложка к книге E. Imber-Black & J. Roberts, 1992).
Основным записывающим устройством, используемым в психофизиологических исследованиях, является полиграф – прибор, позволяющий одновременно фиксировать изменения электрического потенциала по нескольким каналам (2, 4, 6, 8, 16 и более) (рис. 2.2). Если электроды накладывают на поверхность головы, то получаемая запись называется электроэнцефалограммой (ЭЭГ); если они размещаются на обеих руках, или на руке и ноге, или в области сердца, то запись носит название электрокардиограммы (ЭКГ); в случае, когда они располагаются на тыльной и ладонной поверхности руки, записывается кожно-гальваническая реакция (КГР); при расположении электродов вдоль мышцы можно зафиксировать электрическую активность мышц, т. е. получить электромиограмму (ЭМГ); электроды, расположенные по обе стороны глаз, дают электроокуло грамму (ЭОГ). Более подробно об этих устройствах и методах их применения сказано ниже.
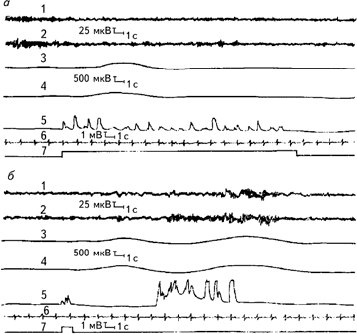
Рис. 2.2. Образец многоканальной записи физиологических функций испытуемого при прослушивании (а) и воспроизведении (б) списка слов. 1, 2 – ЭЭГ, отведения О1 и О2 левое и правое полушария соответственно; 3, 4 – КГР левой и правой руки; 5 – фонограмма (сигналы появляются в моменты предъявления слов магнитофоном или при устных ответах испытуемого); 6 – ЭКГ; 7 – отметка программы опыта (Леутин, Николаева, 1988).
В зависимости от способа наложения электродов и от типа самих электродов при регистрации ЭЭГ, кроме самой ЭЭГ, можно записать также ЭМГ, если человек в процессе обследования будет напрягать лоб или говорить; ЭОГ – если он будет двигать глазами; и даже ЭКГ – если электроды будут, например, сильно прижаты к поверхности кожи и пережмут сосуд, расположенный под ним. Эти ненужные в данный момент для исследователя записи являются помехами и называются артефактами. Электрические сигналы могут генерироваться в некоторых случаях и самим оборудованием, что приводит к появлению наводки. Получение безартефактной записи в психофизиологии сродни искусству и требует многих навыков от исследователя.
В настоящее время существуют специальные программы, позволяющие вычленять безартефактные участки записи, но тем не менее каждую запись рекомендуется предварительно просмотреть, поскольку ни одна программа не сможет пока заменить глаз опытного психофизиолога.
Если раньше исследователи просто изучали характер кривых, то сейчас полиграф обычно сопрягается с компьютером, оснащенным соответствующим программным обеспечением.
При наложении двух электродов на исследуемый участок ткани можно оценить изменение разности потенциала между этими двумя точками. Такую запись называют биполярной. В том случае, когда один из электродов расположен на активной ткани (этот электрод называется активным), а другой (референтный) – на относительно неактивной ткани, запись будет называться монополярной.
Близкородственные науки – психофизиология и физиологическая психология – пользуются полиграфом в различных экспериментальных условиях. Преимущество психофизиологии заключается в том, что в ней оценивают реакции человека без хирургического вмешательства. Это преимущество одновременно является и недостатком, поскольку лишает исследователя возможности непосредственного сопоставления психологических реакций с изменениями активности отдельных нейронов. Физиологическая психология, напротив, активно использует методы внедрения в мозг и другие ткани, и потому в таких экспериментах участвуют либо животные, либо люди, воздействие на мозг которых делают по медицинским показаниям. Это ограничивает интерпретацию такого рода исследования и распространение его результатов на здорового человека. В настоящем учебнике использованы результаты, полученные исследователями обеих областей знания, поскольку они углубляют и дополняют друг друга.
Электроэнцефалограмма и методы ее регистрации
В 1929 г. австрийский психиатр Ганс Бергер обнаружил, что с помощью игольчатых платиновых электродов, помещенных на различные точки поверхности головы человека, можно зарегистрировать электрическую активность мозга. Эти записи он и назвал электроэнцефалограммой (ЭЭГ). Хотя ЭЭГ снимается с поверхности головы, Г. Бергер сумел доказать, что часть электрической активности обусловлена деятельностью мозга, а не покрывающих его поверхность тканей (Berger, 1929).
Открытие Г. Бергера было встречено весьма холодно, и этот метод получил признание лишь после того, как Е. Д. Эдриан и Б. Х. Мэттьюз смогли непосредственно продемонстрировать его на заседании Английского физиологического общества в 1935 г. Сам Е. Д. Эдриан был в качестве испытуемого и, закрывая глаза, показал появление альфа-ритма на ЭЭГ (Хэссет, 1981).
Интерпретация электроэнцефалографических данных требует учета целого ряда параметров – формы волн, их амплитуды, расположения электродов, с которых получена запись, и т. д. Наиболее важными из них являются амплитуда волн и частота. Амплитуда измеряется как расстояние от базовой линии до пика волны. Однако существуют значительные трудности в определении базовой линии, поэтому на практике обычно используют измерение от пика до пика (рис. 2.3). Под частотой понимается число полных циклов, совершаемых волной за 1 сек. Этот показатель измеряется в герцах (Гц). Величина, обратная частоте, называется периодом волны. Известно, что амплитуда волн ЭЭГ находится в обратно пропорциональной зависимости от их частоты (Cooper e. a., 1980).







