


Полная версия
Половой отбор: теория или миф? Полевая зоология против кабинетного знания
Нетрудно видеть, насколько расплывчаты используемые здесь понятия «потери» и «выигрыш». Очевидно, им едва ли можно дать четкое определение. Вот что сказано в одной работе, где речь идет об оценке стоимости так называемых «честных» сигналов. «Операциональные суждения о стратегической стоимости (strategic cost) будут спорными до тех пор, пока не существует строгой методологии ее отделения от эффективной стоимости (efficacy cost). По той же причине не информативно понятие сбалансированные затраты (equilibrium cost)… Только измерение пограничных значений потерь (costs) и выигрыша (benefits) при трансляции сигнала может говорить о том, действительно ли его честность (honesty) поддерживается за счет неких затрат» (Szamado, 2011: 8; курсив мой – Е.П.). Здесь снова уместно повторить слова Р. Левонтина (1978: 25), уже цитированные во введении: «Если переменные состояний или параметры, на которых построена теория, измерить невозможно или если измерение сопряжено с такими ошибками, что невозможно сделать выбор между альтернативными гипотезами, теория становится пустым упражнением в формальной логике, никак не связанным с реальным миром» (курсив мой – Е.П.).
V. Послебрачный половой отбор. Чтобы половой отбор работал в каждом поколении, требуется закрепление репродуктивного успеха преуспевающих самцов. Для этого необходимо существование каких-то механизмов, «страхующих» благоприятную судьбу генов данного конкретного самца. К их числу относят самые разные приспособления. Они могут определяться, например, активностью самца, охраняющего самку от посягательств соперников после копуляции с ней. Пассивными средствами, как полагают, оказывается, в частности, формирование во влагалище самки посткопуляционной пробки. Но особое значение придают здесь явлению «конкуренции спермы», в которой самец может выиграть за счет увеличения числа актов спаривания с данной самкой (см. раздел 10.1).
Таков, вкратце, перечень того спектра явлений, которые сегодня относят к сфере действия полового отбора у животных в той его трактовке, которая идет от Дарвина. Этот спектр уже здесь оказался настолько широким, что покрывает собой не только все, относящееся к сфере биологии размножения, но также ряд вопросов, далеко выходящих за эти рамки – например, касающихся механизмов онтогенеза (флюктуирующая асимметрия) и эволюции отношений между паразитами и их хозяевами. Однако, апологеты «теории» не остановились на этом и попытались модернизировать ее таким образом, чтобы включить в ее сферу также происходящее в мире растений (см. ниже, 1.3). Но, как я уже упоминал, когда теория претендует на объяснение всего без исключения, трудно ожидать от нее четких ответов на конкретные вопросы более узкого содержания. Особенно если основополагающие понятия, такие, например, как «потери» и «выигрыш», предельно аморфны, не поддаются сколько-нибудь строгим оценкам, но могут быть с успехом адресованы обыденному здравому смыслу.
1.2. Поддержка «теории» со стороны абстрактных математических и генетических моделей
До сих пор речь шла о тех вопросах, которые можно пытаться более или менее убедительно обосновать эмпирическими данными. Теперь я остановлюсь на построениях кабинетных теоретиков, которые принято считать формальным доказательством теории полового отбора. Как уже было сказано, их основа была заложена математиком Рональдом Фишером, предложившим схему так называемого «неудержимого процесса полового отбора» (runaway process), имеющего постоянный направленный характер. Саму идею в вольной словесной форме он высказал в своей книге «Генетическая теория полового отбора» (Fisher, 1930), где он, по его словам, «рискнул внести свое в понимание в эволюцию психических явлений, именно вкусов самок (female taste) на основе того же самого процесса селекции»[23] (имеется в виду естественный отбор Дарвина).
Вскоре после выхода книги Фишер получил письмо от внука Ч. Дарвина, Чарлза Гальтона Дарвина[24], в котором тот писал, что он «пока что не понял, как может работать постулируемый неудержимый процесс полового отбора, приводящий экспоненциальному сопряженному усилению признаков самцов и предпочтений к ним самок, которые параллельно нарастают в геометрической прогрессии» (Fisher, 2000: 305–306). Ч.Г. Дарвин спрашивал, нельзя ли отобразить ход этого процесса математически.
В нескольких ответных письмах Фишер изложил свою схему процесса в виде последовательности алгебраических формул. При этом он опирался на пример из орнитологии, взятый им из книги Силоуса «Реальная жизнь птиц» (Selous, 1927). Речь шла о куликах турухтанах (Phylomachus pugnax), а не о павлинах, как обычно пишут, ссылаясь на построения Фишера[25]. «Силоус наблюдал, – писал тот, – как самка важно (with perfect self composure) расхаживает в толпе самцов[26], но не торопится выбрать кого-нибудь из них. Это замечательный экологический пример (perfect ecological framework). Самка выбирает стиль (fashion) орнамента своих будущих сыновей» (там же: 308).
Модель «неудержимого процесса полового отбора» Р. Фишера. Чтобы показать, каким способом этот автор попытался внести свой вклад в «эволюцию психических явлений», я буду вынужден привести текст одного из писем Фишера Ч.Г. Дарвину (25 октября 1932 г.) почти полностью. Надеюсь, что эти материалы помогут рассеять миф, согласно которому Фишер «объяснил» ход процессов полового отбора.
«Обозначим красоту (beauty) самца через х, а вкусы (taste) самки через у. Эти [признаки – Е.П.]. будут варьировать вокруг некоторых средних значений хср.и уср. Из них на хср пока что можно не обращать внимания, поскольку в природе для этой величины нет нулевого значения. Что касается уср то у него нулевое значение есть: уср = 0 обозначает полную индифферентность самки, а уср показывает среднюю интенсивность ее предпочтений. Мы можем допустить для удобства, что х и у определяются генетически, так что их средние значения у потомков пары равны среднему значению для их родителей и что для каждого значения шкала выбрана таким образом, что средние значения как (х – хср)2, так и (у – уср)2 равны единице. Они могут быть скоррелированы в той степени, которую мы и должны выяснить, так что мы можем заменить (х-хср) (у – уср) символом г.
Мы можем допустить, что красоту (beauty) удается измерить объективно – например, взяв длину перьев самца турухтана[27], тогда как вкусы (taste) самок могут быть измерены (measured) через ее реальные действия (actual performance). Самка, не обладающая такими вкусами, будет спариваться наугад (at random), то есть со средними значениями для исхода нескольких случаев (a number of trials), так что средняя величина по самцам, ставшим ее партнерами, будет хср. Избирательная самка избежит некоторых возможностей спариться с некрасивыми (ugly) самцами и тем самым превысит среднее значение этой переменной (will score a higher average). Имея в виду нашу шкалу измерений, можно сказать, что показатель такой самки есть у, если красота избранного ей самца составляет хср + kу. Здесь k есть величина (datum), зависящая от силы способности к распознаванию (powers of discrimination), реальной ситуации при выборе, предпосылок к его осуществлению (opportunities for choice) и т. д. (etc).
Если самец со свойством (specifi cation) х1,у1 спарится с самкой x2y2, показатели их потомков будут варьировать вокруг среднего (x1 + х2)/2, (y1 + у2)/2. Единственная гипотеза относительно наследования, в которой мы нуждаемся, состоит в том, что в потомстве этих особей х и у не скоррелированы. Если это так; то среднее г для потомков особей этой генерации будет ни чем иным как
(x1 – xср + х2 – xср)(y1 – yср + у2 – yср)/4
усредненное по всем спариваниям (averaged over all matings). Если здесь мы видим то же, что и в предыдущем поколении, то получается, что r для среднего
(x1 – xср)(y1 – yср) = r и (х2 – xср)(у2 – yср) = r
в то время как для прочих (while for the rest)
(x1 – xср)(у2 – yср)= ky2(у2 – yср) = k и (x2 – xср)(у1 – yср) = r2k
как это следует из усреднения по самцам, спарившимся с самкой типа x2,у2.
В таком случае отсюда следует
2r = k(l + r2) или r = (1 – √1 – k2)/k[28]
и что отбор, который в каждом поколении повышает усреднение по x на kyср/2, должен увеличивать усреднение по y на kryср/2, то есть yср растет в геометрической прогрессии (при том допущении, что k, и, следовательно r, остаются постоянными). Разумеется, я оставляю в стороне все проверки, часть из которых кажется несущественными сегодня, в то время как другие могут оказаться важными позже».
Это письмо Фишер завершает следующими словами: «Сообщите мне, помогли ли Вам мои разъяснения, чтобы продвинуться вперед. Я чувствую себя совершенно неудовлетворенным тем, что не в состоянии привести полную аргументацию (to get the argument across), но надеюсь, что идея, согласно которой х и у должны быть скоррелированы, сможет устранить те трудности, о которых Вы писали».
О моделях, подобных рассмотренной, с точки зрения эксперта в вопросах популяционной генетики. Через 48 лет после того как научная общественность была осведомлена о том, что идея полового отбора, высказанная Ч. Дарвином в 1871 г., получила «окончательное подтверждение» в модели Фишера, выдающийся исследователь в области генетики популяций Р. Левонтин писал: «Несмотря на то, что теория популяционной генетики хорошо разработана, ее пока еще можно считать теорией лишь в первом приближении. Она все еще зависит от огромного числа упрощающих допущений и примитивных моделей, так что противоречия между наблюдениями и теоретическими предсказаниями могут пока что вскрыть лишь очевидную поверхностность ее предсказаний» (Левонтин, 1978: 237; курсив мой – Е.П.).
Нетрудно согласится с этим автором в том отношении, что «еще задолго до появления прямых доказательств научные работники выносят на обсуждение свои глубоко укоренившиеся предубеждения, и чем важнее предмет спора, чем сомнительнее доказательства, тем глубже предубеждения…» (там же: 37). Наконец, в преамбуле к разделу своей книги под рубрикой «Теория», Левонтин так оценивает роль последней в первой половине XX столетия: «В течение многих лет популяционная генетика была необычайно обширной и мощной теорией, лишенной, в буквальном смысле слова, фактов, которыми она могла бы оперировать. Ее можно было сравнить со сложной и превосходной машиной, предназначенной для переработки сырья, которое никому не удается добыть. Время от времени какой-нибудь необыкновенно удачливый или опытный изыскатель наталкивался на природную жилу богатой руды, и тогда часть механизмов пускали в ход, чтобы убедить тех, кто финансирует машину, что она и в самом деле может действовать. Но большую часть времени машина оставалась в руках изобретателей, которые вечно ее ремонтировали и совершенствовали в ожидании того дня, когда ее можно будет запустить на полную мощь и начать получать продукцию» (там же: 194).
На мой взгляд, нечто подобное происходит, когда теоретики, один за другим, вновь и вновь предпринимают попытки модернизировать модель Фишера, вводя в ее основания какие-либо новые переменные[29]. К сожалению, от результатов этой деятельности трудно ожидать глубокого понимания процессов, происходящих в эволюции органического мира. Каждому здравомыслящему биологу должно быть совершенно очевидно, что схема Фишера не может быть, в принципе, приближена к этой реальности. Она вообще не имеет ничего общего не только с биологией вообще, но даже с начальными представлениями о генетике. Возьмем хотя бы исходную посылку, на которой строятся все последующие умозаключения алгебраического характера: «Обозначим красоту самца через х, а вкусы самки через у». О каких усредненных значениях этих категорий, под каждым из которых кроется сложнейшая система механизмов и корреляций, может идти речь? А ведь все дальнейшие построения Фишера базируются именно на оперировании такого рода средними значениями.
Критика модели Фишера с позиций современной количественной генетики.
В статье Домини (Dominey, 1983) были высказаны сомнения относительно соответствия теории Фишера принципам этой дисциплины. Автор статьи кратко пересказывает идею Фишера, ссылаясь на второе издание его книги (Fisher, 1958). Он пишет: «“Неудержимый процесс”, как его именует Фишер, берет начало в ситуации, когда признак (trait) самца имеет некоторые преимущества, вне зависимости от тех или иных предпочтений самок. Коль скоро эти особенности самцов, как и выбор их самками по этому признаку, должны быть селективно выгодны, частоты того и другого будут нарастать в популяции. В конце концов, по мере того, как этот процесс прогрессирует, самки, которые выбирают самцов, лишенных рассматриваемых характеристик, должны оказаться в проигрыше (disadvantage) просто потому, что их сыновья не смогут приобретать такие качества и, таким образом, утратят возможность быть привлекательными для самок. Но признак самцов может продолжить свое развитие из-за сохраняющихся предпочтений самок, даже в том случае, если он утратил свое первоначальное преимущество, став чересчур гипертрофированным и вредным с точки зрения естественного отбора. Фишер утверждает, что скорость процесса должна нарастать в геометрической прогрессии в силу развития генетической взаимозависимости между крайне выраженными предпочтениями самок и максимальной выраженности признака у самцов.
В основе всей концепции лежит допущение, что признак самца должен быть наследуемым, то есть здесь должна иметь место аддитивная генетическая дисперсия[30], а следовательно, и корреляция между фенотипами отца и его сыновей. При отсутствии такой корреляции самка, выбирающая самцов с экстремально выраженным признаком, едва ли сможет производить больше сыновей того же типа, нежели самка, спаривающаяся наугад. Поскольку фенотипы сыновей самки не будут соответствовать фенотипу их отцов, она утратит все преимущества от выбора самцов с экстремальной наружностью (в плане воспроизводства потомства с требуемым фенотипом) и, вероятно, потерпит всевозможные затраты при поисках партнера (время, энергию и пр.). В этой точке предпочтения самок должны вступить в стадию редукции. Эта трудность для построений Фишера была четко обозначена Мейнардом Смитом (Maynard Smith, 1978: 172).
Таким образом, построения Фишера должны быть существенно зависимы от фактора аддитивной генетической дисперсии. Но, согласно предсказаниям современной количественной генетики, при постоянном направленном отборе некоего признака должно происходить уменьшение аддитивной генетической дисперсии и, в конечном итоге, полная утрата[31]. Такая утрата может быть обусловлена фиксацией отбираемых аллелей или, в случае сверхдоминирования в данном локусе, результатом будет формирование устойчивого полиморфизма. В последнем случае в потомстве самок, спаривающихся наугад (без выбора самцов с генотипом, соответствующих ее собственному) фенотипы ее сыновей будут включать в себя разное соотношение гомо- и гетерозигот по данному признаку. В случае потери аддитивной генетической дисперсии процесс чрезмерного развития (exaggeration) признака остановится даже в отсутствие противодействия со стороны естественного отбора» (Dominey, 1983: 495–496).
В той же статье много внимания уделено другой мысленной конструкции, столь же охотно принимаемой на веру и еще менее реалистичной, чем построения Фишера и его эпигонов. Я имею в виду так называемый «принцип гандикапа» за авторством А. Захави (Zahavi, 1975)[32]. Об этой идее было упомянуто выше (1.1), но стоит еще раз повторить ее суть. Мерой качества генотипа самца является его способность дожить до размножения вопреки обладанию им структурами, затрудняющими существование. Таковы, например, роскошный хвост павлина, неудобный в полете, или шикарные, но очень тяжелые рога быка-оленя. Такого рода структуры служат, согласно этим представлениям, указанием самке на то, что самец прошел «тест полового отбора», подтвердив тем самым высокое качество своих генов. В том, что самец не скрывает свои пороки (трудности, с которыми ему приходится сталкиваться), но откровенно выставляет их напоказ, и состоит «честность» его коммуникативного поведения. Эти умозрительные построения, рожденные игрой ума в тиши кабинета, едва ли можно воспринимать всерьез.
Приходится только удивляться тому, что такого рода мысленные конструкции, не проверяемые в принципе и потому не относящиеся, строго говоря, к сфере истинной науки, могут серьезно обсуждаться годами. Понятно, что подтвердить их справедливость удается только с применением все тех же умозрительных, отвлеченных моделей. Как пишет А. Графен[33], «… модели из области теории игр показывают, что принцип гандикапа работоспособен: каждый организм максимизирует свою приспособленность, и сигналы следует признать честными (signals are honest)» (Grafen, 1990: 517). На мой взгляд, комментарии излишни.
1.3. «Половой отбор» у растений
«Новое» в «теории полового отбора»[34]. В июне 1993 г. в Сноубирде (Юта, США) состоялся симпозиум по теме “Половой отбор у растений и животных». Его материалы открываются статьей С. Арнольда, который поставил своей задачей сформулировать концепцию полового отбора, применимую не только к животным, но и к растениям (Arnold, 1994). Стоит мимоходом заметить, что ее автор – герпетолог, специалист по биологии саламандр и змей, никогда прежде не замеченный в интересах к проблемам размножения у растений. Резюме этой статьи сводится к следующему. «Статья служит введением к теме симпозиума и направлена на поиски определения полового отбора, которое было бы применимо к происходящему в обоих царствах… Рассмотрены 10 критериев, которые могли бы быть положены в основу такого определения. Я утверждаю, что понятия «выбор полового партнера» (mate choice) и «половая конкуренция» (sexual competition) проблематичны и в определении их следует избегать. Вместо них я предлагаю критерий успеха спариваний (mating success), которое может служить унифицированным понятием (unifying concept), пригодным в отношении как растений, так и животных» (Arnold, 1994: 1; курсив мой – Е.П.).
Автор статьи аргументирует необходимость в дефиниции следующим образом. «Ясно, – пишет он, – что если явление не определено, оно не поддается измерениям[35]. Но если идея полового отбора столь трудна для понимания (so troublesome), почему не отказаться от самого термина? Это означало бы прервать историческую преемственность, отказавшись от представлений Дарвина и от понятия, бывшего в употреблении на протяжении 130 лет. Даже если мы отбросим его, понадобится найти нечто взамен. Когда половой отбор противопоставляется другой селективной силе[36], его обозначение специальным термином особенно полезно. И, наконец, поиски такого определения, которое годилось бы для царств растений и животных, привело бы к интеллектуальному оживлению в обеих научных сферах и повело бы нас в новых направлениях» (Arnold, 1994: 3).
Все эти рассуждения едва ли можно признать логичными. С одной стороны, автор настаивает на том, чтобы изъять из понятия «половой отбор» главное его содержание в трактовке Дарвина (выбор половых партнеров самками и конкуренция между самцами за доступ к половым партнерам). Именно эти явления служили предметом исследований в зоологии, по словам Арольда, «на протяжении 130 лет»[37]. С другой стороны, он предлагает сохранить термин в таком выхолощенном состоянии, чтобы не прерывать историческую традицию. Если бы в науке следовали этому рецепту, мы бы до сих пор думали, что причиной горения служит присутствие флогистона в горючих субстанциях.
Не менее наивным оказывается утверждение автора, что «Чем проще определение, тем лучше» (Arnold, 1994: 9). Посмотрим, каков же итог построений, изложенных в статье. «В свете изложенного я предлагаю следующее определение полового отбора. Половой отбор – это форма селекции, которая проистекает (arises) из различий в успехе спариваний [число половых партнеров (mates), которые произвели[38] потомство на протяжении некоторого стандартизованного отрезка времени]. Это определение выигрывает из-за своей относительной простоты, поскольку лишено попыток дать дефиницию отбору как таковому (??? – Е.П.). В нем я избегаю трудностей применения к растениям категории выбора со стороны самок, а также операциональной дефиниции конкуренции у этих организмов» (там же, курсив автора статьи).
Не хотелось бы подавать читателям пример изощренной схоластики, ведущей в эмпирический тупик. Но я вынужден привести иллюстрацию из обсуждаемой статьи, чтобы показать, как представления ее автора о половом отборе незаметно сводятся к стандартной схеме естественного отбора: приспособление (fitness) есть следствие дифференциальной размножаемости особей (рис. 1.2). Об этом неявно свидетельствует и следующая сентенция самого автора: «Поскольку мое определение выдержано в терминах стоимости приспособленности (fitness currency), его соответствие формальной эволюционной теории самоочевидно (straightforward). Так или иначе, оно лежит в русле сказанного в большинстве произведений Дарвина[39]»
«Половой отбор» у растений: история вопроса. Первая серьезная попытка обосновать существование полового отбору в растительном царстве была предпринята в 1983 г. в книге «Выбор полового партнера у растений» (Willson, Burley, 1983). Указывают, что ее появление повлек за собой бурный рост публикаций на эту тему (http://web.pdx.edu/~cruzan/SexualSelection inPlants.pdf)[40].
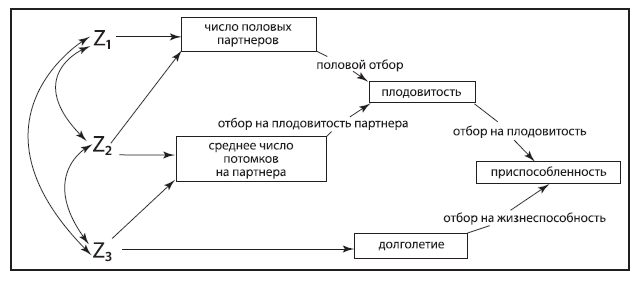
Рис. 1.2. Иерархия компонентов приспособленности, диктуемых разными формами отбора. Z1, Z2, Z3 – три гипотетические фенотипические признака. Стрелки с двумя остриями показывают корреляции между этими признаками. Из: Arnold, 1994.
В основе построений авторов книги лежат два постулата из арсенала социобиологии: родительский вклад и конкуренция полов. Первый из них был выдвинут Р. Трайверсом еще на заре становления социобиологии. Суть понятия родительский вклад состоит в следующем. Это издержки, связанные с вероятностью риска для родителя произвести такое потомство, которое будет недостаточно успешным. Если вероятность этого выше для самок, чем для самцов, селективность первых при выборе потенциальных половых партнеров должна быть достаточно высокой, чтобы избежать ошибки. В таком случае самки оказываются «лимитирующим ресурсом» для самцов, так что конкуренция между ними в половой сфере обостряется. При этом интенсивность полового отбора выражается в соотношении репродуктивного успеха самцов и самок, так что для каждого из родителей он может быть выражен как выигрыш (benefit) для их потомства минус затраты (cost) – именно снижение их количества или качества в будущем (Trivers, 1972).
Конец ознакомительного фрагмента.
Текст предоставлен ООО «ЛитРес».
Прочитайте эту книгу целиком, купив полную легальную версию на ЛитРес.
Безопасно оплатить книгу можно банковской картой Visa, MasterCard, Maestro, со счета мобильного телефона, с платежного терминала, в салоне МТС или Связной, через PayPal, WebMoney, Яндекс.Деньги, QIWI Кошелек, бонусными картами или другим удобным Вам способом.
Примечания
1
В последних трех главах та же идея обсуждается в применении к человеку, но эта тема выходит за пределы тех проблем, которые я буду рассматривать в этой книге.
2
Вот та часть тезауруса, которая в работе Г.П. Дементьева касается вопросов пола: половая система – влияние щитовидной железы на ее функции и ее развитие; стимулирование ее деятельности видом гнезда; половое возбуждение, выражение его в крике; половое развитие, влияние на него тиреоидектомии; половой диморфизм – выражение его: в интенсивности метаболизма, в количестве перьев, в окраске клюва, в размерах клюва, в распределении шпор, в структуре скелета, в форме клюва, в числе рулевых перьев; половой состав птиц; половой цикл самок и самцов, синхронизация его; половые гормоны, действие их: на развитие признаков пола, на рост пера, на физиологические процессы; половые железы – влияние гормона гипофиза на их формирование, значение их гормонов для обмена веществ; трансплантация их кастрированным особям противоположного пола; половые клетки первичные.




