


Полная версия
Логика случая. О природе и происхождении биологической эволюции

Рис. 2–2. Один из пандативов базилики Святого Марка в Венеции. Фото Марии Шнитцмейер, Викисклад.
В более ранней статье по сходной тематике Франсуа Жакоб (один из первооткрывателей регуляции генов и автор нескольких других плодотворных идей в бактериальной генетике, см. гл. 5) ввел метафору мастера-самоучки. Отталкиваясь прежде всего от сравнительного анализа механизмов развития, Жакоб положил в основу своих рассуждений, что эволюция действует не как инженер или дизайнер, а скорее как ремесленник-самоучка, причем чрезвычайно зависимый от предыдущего опыта при решении стоящих перед ним проблем: «Сложно проводить аналогии между естественным отбором и какими-то аспектами человеческого поведения. Однако если очень хочется поиграть в сравнения, то можно сказать, что естественный отбор работает не как изобретатель или инженер. Он работает как дилетант – мастер на все руки, который не знает точно, что он собирается создать, и при этом использует все, что подвернется под руку, будь то обрывки ниток, куски дерева или старые коробки; короче, он действует как тот мастеровой, который использует все, что есть в его распоряжении, чтобы сделать хоть что-то, лишь бы работало» (Jacob, 1977).
Ключевым выводом концепции ремесленника-самоучки становится то, что итоговый результат эволюции непредсказуем, или по крайней мере его невозможно предсказать, не зная в деталях всех предшествующих событий. Другими словами, если взять и «проиграть эволюционную пластинку заново» (любимая метафора Гулда) в некоем мысленном эксперименте, то результат будет отличен от того, что мы наблюдаем в реальности, возможно до неузнаваемости; мы вернемся к этому обсуждению позднее в этой книге (см. гл. 13).
Эволюция в мире микробов и вирусов и трехдоменное древо жизни
Вероятно, в ходе развития биологии наибольшее влияние на изменение представления об эволюции оказало распространение эволюционных исследований на мир микробов, а именно одноклеточных эукариот (протист), прокариот (бактерий и архей) и вирусов. Дарвиновское представление об эволюции и все достижения эволюционной биологии нескольких последующих десятилетий базировались исключительно на исследовании животных и растений, тогда как одноклеточные эукариоты (протисты) и бактерии (монеры) были сугубо номинально размещены у корня древа жизни Эрнстом Геккелем и его последователями. Хотя к 1950-м годам генетический анализ бактериофагов и бактерий продвинулся настолько, что стало очевидным, что эти формы жизни обладают эволюционирующими геномами, СТЭ не принимала во внимание эти открытия. То, что бактерии (не говоря уже о вирусах) эволюционируют по тому же самому принципу и с использованием тех же механизмов, что и животные и растения, отнюдь не очевидно, учитывая все их разительные биологические отличия от многоклеточных организмов, и в особенности из-за отсутствия у них типичного полового размножения и репродуктивной изоляции, ключевых для видообразования среди животных и растений.
Фактически прокариоты стали «видны» эволюционным биологам в 1977 году, после выхода революционной работы Вёзе и его коллег по филогенезу рРНК (Woese, 1987)[26]. Рассмотренное в общем контексте, открытие Вёзе является эпохально важным и, возможно, даже заслуживает сравнения с открытием структуры ДНК. Вёзе установил, что в одной молекулярной структуре, а именно последовательности нуклеотидов рРНК, выявляется очевидная консервативность во всем диапазоне клеточных форм жизни. Кроме того, чрезвычайно информативным оказался и филогенетический анализ этой универсальной консервативной молекулы: он показал, что рРНК, в некотором приближении, эволюционирует с постоянной скоростью, то есть подчиняется модели молекулярных часов. Это привело к еще одному важному открытию, ставшему одним из символов эволюционной биологии конца ХХ века, – трехдоменному древу жизни (см. рис. 2–3; Woese et al., 1990). Тремя доменами являются бактерии, археи и эукариоты. Домен архей был открыт Джорджем Фоксом и Вёзе сравнительным анализом рРНК, когда в новой группе ничем, казалось бы, не примечательных «бактерий» обнаружились существенные отличия как от остальных бактерий, так и от более сложных эукариотических организмов. В дополнение к разграничению трех доменов, Вёзе и его коллеги использовали филогенетический анализ рРНК для идентификации нескольких основных ветвей архей и бактерий (Woese, 1987). Из этого следовало, что эволюция прокариот столь же доступна для изучения, как эволюция сложных эукариот, – концепция, чуждая микробиологам до работы Вёзе (Stanier and Van Niel, 1962). Благодаря достижениям Вёзе, его сотрудников и последователей появилась все усиливающаяся тенденция приравнивать филогенетическое древо рРНК, с его трехдоменной структурой, к древу жизни Дарвина и Геккеля (Pace, 2009а, 2006). В течение нескольких лет после публикации открытий Вёзе стало ясно, что топологически древо рРНК (по крайней мере, в своих основных чертах) конгруэнтно деревьям некоторых из самых консервативных белков, таких как рибосомные белки, факторы трансляции, субъ единицы ДНК-зависимой РНК-полимеразы и мембранные АТФазы.
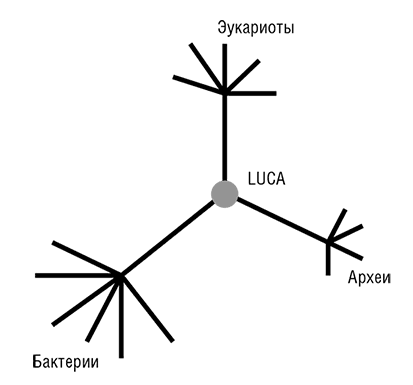
Рис. 2–3. Трехдоменное древо жизни Вёзе.
Две группы исследователей независимо друг от друга пришли к блестящей идее о том, как определить положение корня в эволюционном дереве, которое до этого было бескорневым (рис. 2–3). Для этой цели можно использовать древние паралоги, которые представлены в (почти) всех организмах и, таким образом, можно с уверенностью заключить, возникли в результате дупликации, предшествующей последнему общему предку всех живых организмов (LUCA). Когда дерево строится совместно для двух паралогичных множеств древних ортологов, положение корня между ними определено однозначно, и таким образом корень может быть выведен для каждого из множеств ортологов (см. рис. 2–4; Gogarten et al., 1989; Iwabe et al., 1989). Результаты анализа двух пар древних паралогов, факторов трансляции и субъединиц мембранных АТФаз были полностью совместимы и поместили корень на бактериальную ветвь, установив таким образом кладу архей-эукариотов (см. рис. 2–4). Тем не менее даже в догеномную эпоху было ясно, что не все деревья белок-кодирующих генов имеют ту же топологию, что и дерево рРНК; причины этих отличий оставались неясными и, как предполагалось, включали (за исключением артефактов метода) горизонтальный перенос генов (ГПГ. Smith et al., 1992). Эти расхождения оставались лишь интересным дополнением к трехдоменному ДЖ, но все резко изменилось с наступлением эры геномики.
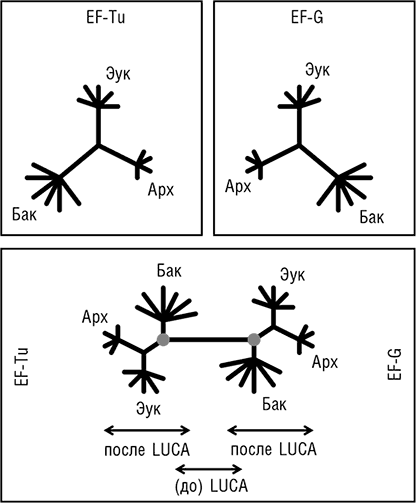
Рис. 2–4. Определение положения корня в трехдоменном древе жизни с помощью древних паралогов. Схематически показаны филогенетические деревья двух широко распространенных факторов инициации трансляции EF-Tu и EF-G, реконструированные независимо (верхние диаграммы) и совместно (нижняя диаграмма). Кружками обозначено вычисленное положение корня в каждом из двух деревьев.
Вирусы и рождение эволюционной геномики
Эволюционная геномика родилась более чем за десять лет до исторического заявления о секвенировании первого бактериального генома. С меньшей помпой (но и не в безвестности) было секвенировано несколько небольших (в рамках 4—100 Кб) различных вирусных геномов, и были разработаны принципы сравнения геномов, наряду с практическими вычислительными методами. Вирусы являются облигатными внутриклеточными паразитами, и вирусные геномы намного меньше и качественно отличаются от геномов клеточных форм жизни. Вирусы обычно не лишены некоторых классов генов, вездесущих и незаменимых в клеточных организмах, таких как гены компонентов системы трансляции и биогенеза мембран. Тем не менее вирусы следуют своей собственной «биологической стратегии» и кодируют все субъединицы вириона, а также по крайней мере некоторые белки, участвующие в репликации вирусного генома. (Одна из центральных тем этой книги – ключевая роль вирусов в эволюции биосферы, так что я возвращаюсь к ней достаточно подробно в гл. 10[27].) Несмотря на быструю эволюцию последовательности генома, характерную для вирусов (в особенности вирусов с РНК-геномом), эти ранние сравнительные геномные исследования успешно выявили множества генов, консервативных в больших группах вирусов (Koonin and Dolja, 1993). Возможность структурного и функционального картирования всего генома определенной формы жизни была реализована в этих исследованиях впервые, и это стало краеугольным камнем эволюционной геномики. Кроме того, было сделано непредвиденное и важное обобщение: в то время как некоторые гены консервативны для удивительно обширного разнообразия вирусов, архитектура генома, структура вириона и биологические свойства вирусов демонстрируют гораздо большую пластичность (см. гл. 5 и 10, где этот вопрос обсуждается подробнее).
Эндосимбиоз
Гипотеза о том, что некоторые органеллы эукариотических клеток, в частности хлоропласты растений, произошли от бактерий, не намного моложе «Происхождения…» Дарвина: некоторые исследователи высказали эту идею в конце XIX века на основе микроскопического исследования клеток растений, показавшего заметное структурное сходство между хлоропластами и цианобактериями (известными тогда как сине-зеленые водоросли). Концепция симбиогенетической эволюции была последовательно представлена Константином Мережковским в начале XX века[28]
Конец ознакомительного фрагмента.
Текст предоставлен ООО «ЛитРес».
Прочитайте эту книгу целиком, купив полную легальную версию на ЛитРес.
Безопасно оплатить книгу можно банковской картой Visa, MasterCard, Maestro, со счета мобильного телефона, с платежного терминала, в салоне МТС или Связной, через PayPal, WebMoney, Яндекс.Деньги, QIWI Кошелек, бонусными картами или другим удобным Вам способом.
Примечания
1
Перевод заглавия этого введения представил серьезные трудности. В английском оригинале было towards a postmodern synthesis. Это конечно же игра слов: с одной стороны, postmodern означает просто «после Modern Synthesis» (то, что в русской литературе обычно называется синтетической теорией эволюции, СТЭ), а с другой – «постмодернистский». Как это передать по-русски, совершенно не очевидно. Хуже того, этот нехитрый каламбур неоднократно обыгрывается в дальнейших главах. Никакого способа справиться с этой трудностью, кроме написания этого примечания, ни переводчикам, ни автору в голову не пришло (примеч. авт. к русскому изданию здесь и далее курсивом).
2
Во многом эти представления опираются на публикации крупнейшего современного эволюциониста Форда Дулитла, которые цитируются в соответствующих главах.
3
По иронии судьбы, magnum opus Ламарка «Философия зоологии» был напечатан в год рождения Дарвина.
4
Удивительно точное и глубокое описание непосредственного воздействия и публичного признания книги Дарвина можно найти в романе Джона Фаулза «Любовница французского лейтенанта».
5
Естественно, сам Дарвин не использовал термин «дарвинизм»; этот не очень удачный и едва ли дальновидный неологизм введен последователем и защитником Дарвина, Томасом Генри Гексли, в отзыве на «Происхождение…» (Huxley, T. H. 1860. Darwin on the origin of Species. Westminster Review: 541–570). Это слово имеет некоторый оттенок догматичности, если не псевдонаучности, по созвучию с другими хорошо известными «-измами», например марксизмом, или фрейдизмом, или даже лысенкоизмом (по-русски – лысенковщиной; о лысенковщине см. гл. 9). Естественно, что никто не говорит о ньютонизме или эйнштейнизме, а слово «менделизм» (обычно в составе сложных слов «менделизм-вейсманизм» или «менделизм-морганизм») употреблялось исключительно в негативном контексте антинаучно мыслящими последователями Лысенко в Советском Союзе. Тем не менее термин, предложенный Гексли, твердо прижился и даже привлекает своей емкостью. В этой книге я использую его исключительно для описания «первой синтетической теории эволюции», которая была разработана Дарвином в «Происхождении…», а затем усовершенствована в последующих работах Гексли, Уоллеса, Вейсмана, Геккеля и других ранних последователей Дарвина.
6
Выражение «неупрощаемая сложность» было придумано Майклом Бихи, одним из основных сторонников антиэволюционной гипотезы разумного замысла (РЗ), в его (печально) известной книге «Черный ящик Дарвина» (Behe, M. J. 2006. Darwin’s Black Box: The Biochemical Challenge to Evolution. New York: Free Press). Для Бихи и других сторонников РЗ «неупрощаемость» сложных биологических структур является будто бы подтверждением (даже доказательством) неизбежности РЗ. Конечно же РЗ – это злостная чушь, но термин «неупрощаемая сложность» вполне выразителен, хотя эволюционные биологи предпочли бы говорить о «видимой» или «кажущейся» неупрощаемости сложных структур.
7
Посетитель музея Менделя в Брно имеет возможность рассматривать экземпляр немецкого перевода «Происхождения видов…», густо испещренный пометками Менделя. Автор был там уже после публикации английского оригинала этой книги и остался под сильным впечатлением.
8
Сэр Рональд Фишер был настоящим гением. (Fisher Box, J. 1978. R. A. Fisher: The Life of a Scientist. New York: Wiley.) Он был фактическим основателем не только популяционной генетики, но и, во многом, современной статистики и ввел математическое определение информации задолго до Клода Шеннона. В эту книгу мы включили также другие примеры его замечательных научных предвидений. В то же время сэр Рональд посвятил большую часть своей карьеры делу евгеники, области исследования, которая в настоящее время рассматривается как псевдонаука и граничит с преступлением. Следует избегать суждения о великих умах даже относительно недавнего прошлого по сегодняшним меркам.
9
В русской литературе используется термин «синтетическая теория эволюции» (СТЭ), который и употребляется далее в этой книге.
10
Это уже сделано во многих различного уровня учебниках и монографиях, как обзорных, так и глубоко специализированных. Взвешенное, умеренно специализированное представление данной темы можно найти в издании D. L. Hartl and A. G. Clark (2006). Principles of Population Genetics, Sunderland, MA: Sinauer Associates.
11
В принципе если адаптивный ландшафт строится для гена, то число измерений будет равняться количеству нуклеотидных сайтов. Взаимодействие между сайтами (эпистаз) уменьшает размерность.
12
Формально теорема Фишера не запрещает всё нисходящее движение, потому что она касается только той части адаптивных изменений, которые обусловлены отбором. Фишер, однако, считал, что на практике большинство, если не все популяции слишком велики, чтобы то явление, которое Райт обозначил как дрейф, могло иметь какое-либо влияние на них. Это было предметом ожесточенных споров между Фишером и Райтом. Окончательным победителем оказался, конечно, Райт.
13
Эта знаменитая фраза Добржанского является названием его эссе, опубликованного в журнале «Американский учитель биологии» (Dobzhansky, T. 1973. Nothing in biology makes sense except in the light of evolution // The American Biology Teacher 35: 125–129). В целом это поразительный текст. Хотя большая часть эссе удивительно ясно объясняет идею эволюции, заключительные абзацы посвящены выразительной пропаганде совместимости эволюции и христианства, и трудно избавиться от впечатления, что именно это и было главной целью автора. Добржанский, как истинный прихожанин Русской православной церкви, верил, что Бог осуществил свой план сотворения мира как развертывающийся сценарий эволюции жизни. Более того, Добржанский изобретательно клеймит отрицание эволюции как кощунство, так как данная позиция подразумевает, что Бог – мошенник, который намеренно вводит человечество в заблуждение, предоставляя многочисленные доказательства эволюции. Я подозреваю, что не каждый, кто цитирует это изречение Добржанского в дискуссиях о преподавании эволюции, действительно читал это эссе.
14
Иногда та же самая аббревиатура расшифровывается как Last Universal Common Ancestor (последний универсальный общий предок). Однако представляется полезным подчеркнуть, что речь идет именно об общем предке всех клеточных форм жизни. Мы вернемся к этой теме в главе об эволюции вирусов.
15
Сами по себе эти принципы описываются в элементарных учебниках, но рассматриваемый здесь информационно-теоретический подход уже не так тривиален, так что я счел необходимым явно перечислить эти принципы.
16
Этот принцип может быть подвергнут достаточно обоснованному сомнению в свете открытия разнообразных явлений эпигенетической наследственности (в гл. 9 мы коснемся этих явлений, хотя и слишком кратко). В данной формулировке вся эпигенетика спрятана в слове «косвенно», но автор отдает себе отчет в спорности такого подхода.
17
Чаргафф не оценил вовремя исключительной важности своего открытия, и тот факт, что двум высокомерным молодым людям, не знавшим химии, удалось открыть тайну жизни, которой он, эксперт-химик, не понял, исполнил Чаргаффа горечи до конца его долгой жизни, вдохновляя его едкие, чтобы не сказать язвительные, книги. (Chargaff E. Heraclitean Fire: Sketches from a Life Before Nature. New York: Rockefeller University Press, 1978.)
18
Строго говоря, это верно лишь для идеализированных систем репликаторов, размножающихся без ограничений, то есть экспоненциально. Именно такие системы рассматривал Дарвин вслед за Мальтусом. В более реалистических системах с ограниченным (параболическим) размножением наблюдается «выживание всех», а не только наиболее приспособленных, и для селекции и дрейфа необходимы дополнительные условия, такие как компартментализация (Szathmary E. The origin of replicators and reproducers. Philos Trans R Soc Lond B Biol Sci. 2006 Oct 29;361(1474):1761-76).
19
Не совсем ясно, кто первым сформулировал принцип, который я здесь называю ПОР. Говорится о том, что в 1930-х годах эти идеи высказывали выдающиеся русские генетики Николай Константинович Кольцов и Николай Владимирович Тимофеев-Ресовский (Шноль С. Э. Герои, злодеи, конформисты российской науки. 2-е изд., дополненное. М.: Крон-пресс, 2001), но я ничего не знаю об их формальной публикации. Эрвин Шредингер приближается к этой идее в знаменитой книге (Шредингер Э. Что такое жизнь? Физический аспект живой клетки / Пер. с англ. 2-е изд. Ижевск: РХД, 2002), но все же не приходит к конкретной формулировке. В англоязычной литературе Ричард Докинз четко формулирует идею в 1976 году в ставшем классикой «Эгоистичном гене» (Докинз Р. Эгоистичный ген / Пер. с англ. М.: Мир, 1993). С другой, абстрактной точки зрения, теория самовоспроизводящихся автоматов, не привязанных ни к какому физическому воплощению, была разработана великим математиком Джоном фон Нейманом (Нейман Дж. фон. Теория самовоспроизводящихся автоматов / Пер. с англ. М.: Мир, 1971).
20
Известная русская поговорка «Не до жиру, быть бы живу» выражает ту же идею упрощенно, но достаточно точно.
21
Здесь уместно сказать несколько слов о метафоре в биологии, тем более что метафора «эгоистичного гена» была отмечена как особенно часто вводящая в заблуждение (Ball, 2011). Вне всякого сомнения, использовать и в особенности интерпретировать эту метафору следует с великой осторожностью, и даже легкого намека на очеловечивание генного «эгоизма» следует избегать всеми силами. Тем не менее я думаю, что метафоры необходимы для развития науки, и покуда научные результаты представляются посредством естественного языка (а не только математических выражений), метафоры неизбежны. Кроме того, хорошая метафора – краткая, запоминающаяся и запечатлевшая важные общие тенденции в широком поле наблюдения – способна сильно подтолкнуть свежее мышление и исследование. Я, например, считаю «эгоистичный ген», «мусорную ДНК» и «адаптивный ландшафт» прекрасными метафорами.
22
Во времена, когда еще не было полных геномных последовательностей, предвидение динамического генома было, вероятно, лучше всего представлено в исчерпывающей монографии выдающегося российского генетика Романа Бениаминовича Хесина (Хесин Р. Б. Непостоянство генома. М.: Наука, 1984). Эта пророческая книга была издана незадолго до безвременной кончины Хесина и стала легендарной среди русских биологов. К сожалению, она, по-видимому, не оказала большого влияния за пределами России.
23
Коли уж мы обратили внимание на причастность сэра Рональд Фишера к евгенике, было бы несправедливо не упомянуть, что еще один патриарх популяционной генетики, Дж. Б. С. Холдейн, был многолетним членом Британской коммунистической партии и, вероятно, из партийной лояльности в течение многих лет поддерживал лысенковскую лженауку (или, по крайней мере, оставлял за ней презумпцию научности). Холдейн был ученым не меньших масштабов, чем Фишер, одним из последних великих универсалов в истории науки (Dronamraju K. R. Haldane and Modern Biology. Johns Hopkins University Press, 1968). Ему принадлежат не только многочисленные важные результаты в математической генетике (в том числе теория генетического груза), но и большой вклад в область кинетики ферментативных реакций и – вероятно, важнее всего – множество удивительно пророческих идей о разнообразнейших предметах, которые можно найти в его книгах и статьях (мы вернемся к некоторым из них в гл. 10). Холдейном также написаны сотни блестящих популярных очерков обо всех аспектах науки, многие из которых опубликованы в коммунистической газете «Дейли уоркер». В 1950 году Холдейн вышел из коммунистической партии, осознав степень разрушений, причиненных советской генетике Лысенко и его шайкой. Холдейн может послужить ярким примером того, что даже величайшие ученые не должны рассматриваться в отрыве от исторического контекста.
24
Эмиль Цукеркандль указал мне, что в его совместных с Лайнусом Полингом ранних работах по молекулярной эволюции проводится явное различие между гомологами, эволюционировавшими вертикальным путем, и теми, которые эволюционировали путем дупликации, хотя не вводилось никаких специальных терминов для обозначения этих разных классов гомологов. У каждого открытия или концептуального прорыва свои предшественники.
25
«Сказки просто так» – прекрасный сборник детских сказок Редьярда Киплинга (Киплинг Р. Сказки просто так / Пер. с англ. К. Чуковского и Л. Хавкиной. М.: Росмэн-Пресс, 2011). В них происхождение некоторых особенностей морфологии животных, таких как хобот слона и панцирь броненосца, возводится к различным своеобразным случаям. Киплинг, кажется, уже осознавал ошибочность панадаптационизма, хотя его выводы не всегда совпадали с выводами Гулда и Левонтина.
26
Здесь стоит сказать несколько слов о Карле Вёзе, создателе молекулярной филогенетики и первооткрывателе архей, который скончался уже после публикации оригинала этой книги, 30 декабря 2012 года, на 85-м году жизни (Goldenfeld N., Pace NR. Retrospective. Carl R. Woese (1928–2012) // Science. 2013 Feb 8;339(6120):661). Конечно, и при его жизни у коллег не было никаких сомнений в том, что он был крупнейшим ученым, подлинным революционером в микробиологии и эволюционной биологии. Однако, по точному выражению Анны Ахматовой, «когда человек умирает, изменяются его портреты». По крайней мере, для автора этой книги теперь очевидно, что в последней трети XX века Вёзе не было равных среди биологов. Его вклад в науку исключительно велик и разнообразен, и конечно же ни в коей мере не сводится к открытию архей. В этой книге его имя упоминается чаще, чем любое другое: во-первых, в связи с его пионерскими исследованиями по эволюции генетического кода, выполненными еще в 60-х годах прошлого века, затем, разумеется, при обсуждении молекулярной филогении и трех доменов клеточной жизни и, наконец, в контексте его глубоких концептуальных работ рубежа двух тысячелетий.
