


Полная версия
Кому мешает ДНК-генеалогия? Ложь, инсинуации, и русофобия в современной российской науке



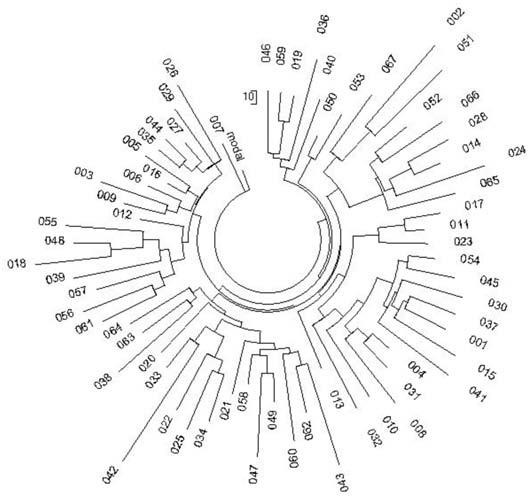
Рис. 1. Дерево из 67 гаплотипов современных немцев гаплогруппы R1a в 25-маркерном формате (Клёсов, 2008)[25].
Если же усреднять поперек всего дерева, то получится, что якобы общий предок всего дерева жил, например, 1200 лет назад (или любое число между 2500 и 600 лет назад). Как видим, этот «общий предок» оказывается фантомным, и такие некорректно полученные результаты искажают выводы работы. К сожалению, так обычно работают популяционные генетики. Именно так, обрабатывая большую серию гаплотипов евреев, они получили якобы возраст якобы общего предка современных евреев (гаплогруппы J1) как 3200 лет назад, и объявили, что это время исхода евреев из Египта (точнее, когда жил Аарон, первосвященник). На самом деле в их исходных данных было две серии гаплотипов, с общими предками 4000 и 1000 лет назад, но авторы работы на ветви гаплотипов не смотрели [26].
Возвращаемся к дереву немецких гаплотипов образца 2008 года (рис. 1). Ниже будет показано, как получаются такие деревья и как их понимать. Здесь отметим, что дерево на рис. 1 показывает общую симметричность (множество мелких ветвей в нем можно анализировать по отдельности, но опыт показывает, что итоговый результат будет практически такой же), и его можно количественно обрабатывать как единую систему, то есть что все 67 гаплотипов происходят от одного общего предка. На все 67 гаплотипов в 12-маркерном формате имелось 208 мутаций от приведенного выше базового гаплотипа, и в 25-маркерном формате – 488 мутаций. В первом случае получаем 208/67/0.02 = 155^183 условных поколений (по 25 лет) от общего предка (напомним, что стрелка указывает на поправку на возвратные мутации), то есть 4575±560 лет до общего предка. Во втором случае, для 25-маркерных гаплотипов, получаем 488/67/0.046 = 158 ^187 условных поколений, то есть 4675±510 лет до общего предка. Как видим, совпадение почти абсолютное для расчетов по 12- или 25-маркерным гаплотипам. Надо сказать, что уже в первом выпуске Вестника, в июне 2008 года, проводились многочисленные расчеты для 37- и 67-маркерных гаплотипов, что тогда было неслыханно. Результаты всех этих расчетов верны до настоящего времени.
Надо сказать, что неопытный читатель тут же запротестует, что, мол, как можно по всей Германии, или любому другому региону судить по каким-то 67 гаплотипам, а там живут миллионы человек. Эта тема была подробно, с экспериментальными данными проанализирована в ДНК-генеалогии, и объяснена в ответе на Вопрос 37, где был задан риторический встречный вопрос – чтобы понять, что море соленое, нужно ли пробовать воду из всех бухточек? А как же химики делают анализ растворов, беря пробу всего лишь в долю миллилитра из ведра, цистерны, озера? Когда делают анализ крови, что, всю кровь из вас выкачивают? Нет. Значит, вопрос о размере пробы поставлен некорректно. Вопрос на самом деле сводится к тому, равномерно ли перемешана система, чтобы по малой пробе, или выборке, судить о всей системе.
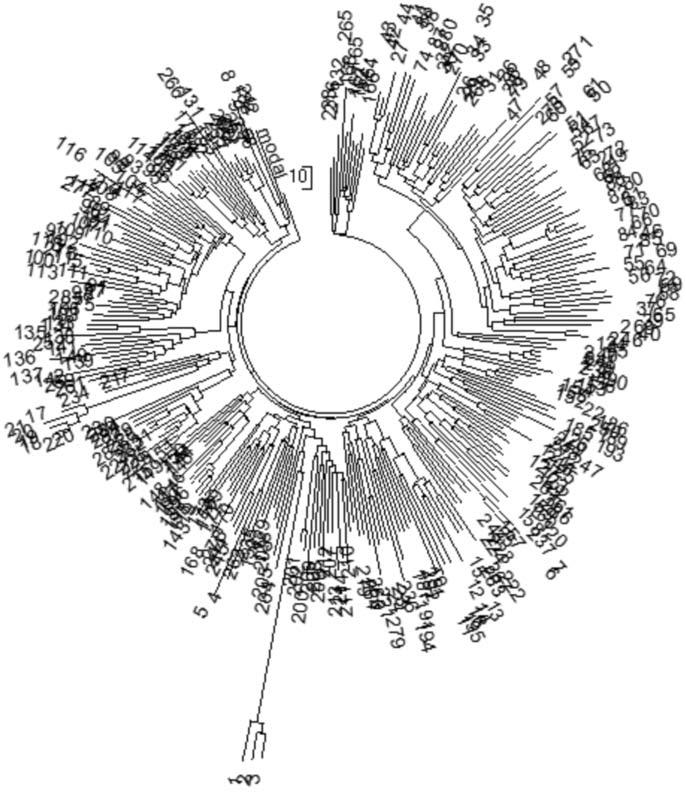

Рис. 2. Дерево из 286 гаплотипов современных немцев гаплогруппы R1a в 67-маркерном формате (база данных IRAKAZ-2015). Три гаплотипа внизу (номера 1, 2, 3) относятся к древнейшему субкладу гаплогруппы, R1a-M420, и в расчеты (см. текст) не включались. Все остальные гаплотипы относились к субкладу М417 (см. диаграмму) и нижестоящим субкладам (на диаграмме не показаны, их несколько десятков). Расчеты проводили на калькуляторе Килина-Клёсова, поскольку ручной расчет системы с более чем 22 тысячи аллелей слишком трудозатратен.
Продолжая это рассмотрение, рассмотрим выборку по Германии уже 2015 года. Теперь в ней 286 гаплотипов в 67- и 65 гаплотипов в 111-маркерном формате (база данных IRAKAZ-2015). Выше – дерево 67-маркерных гаплотипов.
В подписи к рис. 2 для иллюстрации приведена «лесенка» нескольких субкладов гаплогруппы R1a, родительской которых является гаплогруппа R (ее снип – М207, то есть это индекс характерной и уникальной мутации в Y-хромосоме ДНК, которую (мутацию) несут все потомки первопредка гаплогруппы R. Через тысячелетия (в данном случае примерно через 10 тысяч лет) один из потомков первопредка гаплогруппы R получил мутацию М173 (очередной снип), и эта гаплогруппа, которую это потомок основал, став ее патриархом, в классификации называется R1. Она возникла (путем образования мутации М173) примерно 30 тысяч лет назад. Далее, примерно 20 тысяч лет назад, возникла гаплогруппа R1a (путем образования мутации/снипа М420), это произошло, видимо, в Южной Сибири, на Алтае. Три носителя данного снипа были найдены в Европе, в Германии, и они попали на дерево гаплотипов на рис. 6, под номерами 1, 2 и 3. Их последующие потомки – носители субкладов R1a1, R1a1a1, R1a1a, и так далее, на десятки уровней вниз.
Возвращаемся к дереву гаплотипов на рис. 2. Общий предок всех 67-маркерных гаплотипов дерева жил 4848±415 лет назад, всех 67- и 111-маркерных гаплотипов – 4294±298 лет назад, при округлении получаем 4800±400 и 4300±300 лет назад. Среднюю величину можно не брать, и так видно, что она практически равна полученным ранее, в 2008 году, и с совершенно другой выборкой в Германии 4575±560 лет и 4675±510 лет до общего предка, расчитанным по 12- и 25-маркерным гаплотипам.
Таким образом, обе выборки – 2008 и 2014 гг в Германии являются вполне репрезентативными для целей настоящего исследования.
Помимо этого, в ДНК-генеалогии введен принципиально новый метод для расчетов хронологии, основываясь на картине мутаций ДНК- так называемый логарифмический метод. Покажем его эфективность на примере, приведенном выше, для серии из 67 гаплотипов гаплогруппы R1a из Германии (см. рис. 2). В 12-маркерном формате из этой серии всего три гаплотипа идентичны друг другу, значит, это и есть базовые, то есть предковые гаплотипы (хотя для столь малого числа базовых гаплотипов не исключена ошибка в их отнесении к предковым, потому они и называются базовыми, а не буквально предковыми). Получаем: [ln(67/3)]/0.02 = 155 → 183 условных поколений, то есть примерно 4575 лет до общего предка всей серии. Расчет по мутациям, которых в этой серии было 208 (см. выше), дает 208/67/0.02 = 155→183 условных поколений (по 25 лет) от общего предка, то есть 4575±560 лет до общего предка. Совпадение получилось практически абсолютным, но это в данном расчете случайно, потому что чисто статистически в данной серии могли оказаться как 2, так и 4 базовых гаплотипа, и погрешность при таких малых количествах базовых гаплотипов велика. Тем не менее, логарифмический (без подсчета мутаций) и линейный (с подсчетом мутаций) методы расчетов показывают, что серия гаплотипов вполне подходит для расчетов, и что общий их предок действительно жил примерно 4600 лет назад.
Но для 25-маркерных гаплотипов при таком малом числе гаплотипов и таком довольно удаленном общем предке расчет по логарифмическому методу уже невозможен. Даже два базовых 25-маркерных гаплотипа сохраняются всего лишь на протяжении [ln(67/2)]/0.046 = 76 → 83 условных поколений, то есть примерно 2075 лет. После этого и двух базовых гаплотипов не остается. Но даже отсюда можно – просто на взгляд, без расчетов – получить граничную информацию о времени жизни общего предка серии гаплотипов, в данном случае – больше 2100 лет назад. Здесь 0.02 и 0.046 – константы скорости мутаций для 12- и 25-маркерных гаплотипов.
Чем больше по численности серия гаплотипов, происходящих от одного предка, тем точнее расчет времени жизни этого предка. И тем больше экономия времени при расчетах, поскольку логарифмический метод не требует подсчета числа мутаций в гаплотипах. Например, из 2000 12-маркерных гаплотипов субклада R1b-L21 114 гаплотипов являются базовыми, то есть идентичными друг другу. Это дает [ln(2000/114)]/0.02 = 143 → 167 условных поколений, или 4175±570 лет до общего предка. Расчет по мутациям (для 67-маркерных гаплотипов) дает 4325±350 лет до общего предка. Разница между логарифмическим и линейным (то есть по мутациям) методами расчета дает всего лишь 3.6 % разницы, хотя погрешность расчетов ее увеличивает. Если округлить полученные величины с учетом погрешностей до 4200 и 4300 лет до общего предка, разница между ними становится равной всего лишь 2.4 %.
Другой подобный пример – из 2000 12-маркерных гаплотипов субклада RW-Z280 базовыми являются 88 гаплотипов. Мы уже видим, что R1a-Z280 заметно старше, чем R1b-L21 (см. предыдущий абзац). Действительно, это дает [ln(2000/88)]/0.02 = 156 → 184 условных поколений, или 4600±б70 лет до общего предка. Расчет по мутациям (для 67-маркерных гаплотипов) дает 4825±320 лет до общего предка (см. таблицу выше). Разница между логарифмическим и линейным (то есть по мутациям) методами расчета дает всего 4.9 % разницы. Если округлить полученные величины с учетом погрешностей до 4600 и 4800 лет до общего предка, разница между ними становится равной 4.3 %. Для 25-маркерных гаплотипов базовых в серии из 2000 R1a-Z280 остаются только два базовых, что дает [ln(2000/2)]/0.046 = 150 → 176 условных поколений, или примерно 4400 лет до общего предка. Погрешность там велика, поскольку там, где есть только два базовых гаплотипа, легко могли остаться один или три. Но видно, что закономерности в целом соблюдаются.
Как мы видим, мутации в гаплотипах потомков расходятся от предкового гаплотипа как круги по воде, число мутаций легко рассчитывается, и они подчиняются довольно простым количественным закономерностям. Для кругов на воде, расходящихся от места, куда был брошен камень, легко рассчитать, когда был брошен камень, если знать скорость распространения волны и место нахождения круговой волны в данный момент времени. Чем больше прошло времени – тем дальше круги ушли, тем больше они разошлись. Так и в гаплотипах – чем больше время, прошедшее от общего предка, тем больше мутаций накопилось в гаплотипах его потомков. Число этих мутаций связано с временем, прошедшим от общего предка, с числом гаплотипов в серии, и с константой скорости мутации в гаплотипах, и выражается простой формулой: n/N = kt, где n – число мутаций в серии из N гаплотипов, k – константа скорости мутации (в числе мутаций на гаплотип за условное поколение, равное 25 лет), t – число условных поколений, с табличной поправкой на возвратные мутации[27]. На сотнях и тысячах примеров показано, что эта формула работает при любом числе гаплотипов и мутаций в них, и при любом времени, прошедшем от общего предка рассматриваемых гаплотипов.
Однако при очень больших временах, более 10–20 тысяч лет, и особенно более 100 тысяч лет, нужно использовать гаплотипы с «медленными» маркерами, то есть с малыми константами скоростей мутаций, и тем самым снижать число мутаций и число возвратных мутаций. По аналогии, вряд ли целесообразно изучать скорости радиоактивного распада элементов со временами полураспада в тысячелетия, используя секундомер. Или пытаться изучать круги на воде за километры от места, куда был брошен камень, для этого нужно значительно более мощное воздействие. Как всегда, нужен конкретный анализ в конкретной ситуации, единых подходов на все случае жизни не бывает. Варианты конкретного анализа в конкретных ситуациях и рассматривает ДНК-генеалогия. Некоторые ситуации и расчеты мы расмотрим ниже.
Как мы видим, мутации в ДНК-генеалогии – это не только единичные необратимые мутации-снипы, описанные выше, которые определяют гаплогруппы и субклады, но и обратимые мутации, меняющие числа повторов, или аллели, в гаплотипах. В русскоязычной литературе их называют просто «мутации», с пониманием, что это не те мутации (в генах), которые обычно возникают под действием радиации. Переход числа повторов в маркере Y-хромосомы от 25 к 24 или 26 (или наоборот) имеет совершенно другую природу, чем «поломка» гена. Такой переход является следствием спонтанной ошибки ДНК-копирующей «биологической машины», это процесс первого порядка с точки зрения физико-химической кинетики, он не зависит от внешних воздействий. О процессах первого порядка будет рассказано ниже.
Помимо того, что в 2008 году ДНК-генеалогия прошла проверку на реальных (ископаемых) гаплотипах, показав ту же хронологию, в том же 2008 году, в июне, вышел первый выпуск Вестника, который позже стал международным. В первом выпуске были две больших статьи – «Основные правила ДНК-генеалогии» (54 страницы) и «Происхождение евреев с точки зрения ДНК-генеалогии» (179 страниц). С тех пор вышло 73 выпуска Вестника, общим объемом более 15 тысяч страниц, все выпуски в открытом доступе (http://dna-academy.ru/proceedings/, www.anatole-klyosov.com). Со второго выпуска (июль 2008) в разделе «Обращения читателей и персональные случаи ДНК-генеалогии» публикуются ответы на письма читателей из разных стран мира. До сегодняшнего выпуска опубликованы персональные истории в виде ответов на 249 писем читателей. Авторы Вестника – в основном российские специалисты и любители ДНК-генеалогии, но также специалисты из США, Италии, Франции, Словении, Сербии, Италии, Германии, Бельгии, Шотландии и других стран. Помимо этого, в материалах Академии, которая официально утверждена в федеральных органах РФ в июле 2015 года, публикуются персональные интерпретации гаплотипов и снипов для людей, подавших на рассмотрение данные тестирования своих ДНК.
С того же 2008 года опубликованы 25 научных статей по ДНК-генеалогии в академических журналах (21 на английском языке, одна – в старейшем российском академическом журнале «Биохимия»), и три – в журнале «Исторический Формат») вышли 8 книг по ДНК-генеалогии (7 на русском языке и одна на сербском, в Белграде), и 112 научно-популярных статей по ДНК-генеалогии, из них 80 статей на Переформате – сайте, который выпускается профессиональными историками (http://pereformat.ru/klyosov).
Научные статьи в области ДНК-генеалогии публиковались в журналах Human Genetics (2009), J. Gen. Genealogy (2009), Advances in Anthropology (2011–2014), Биохимия (2011), Eur. J. Human Genetics (2014), Frontiers in Genetics (2014), Исторический Формат (2015). Весной 2013 г. автор настоящей книги получил приглашение быть главным редактором журнала Advances in Anthropology («Успехи антропологии»), и сделал его профильным журналом по ДНК-генеалогии (наряду, конечно, с публикациями авторов по разным направлениям антропологии). Первой статьей в «Успехах антропологии» была статья по константам скоростей мутаций в гаплотипах Y-хромосомы и их калибровке, то есть количественной связи с хронологией[28]. Концептуально сделать это было несложно, поскольку кинетика химических и биологических мутаций – прямая специальность автора данной книги, он в свое время заканчивал кафедру академика Н.Н. Семенова, единственного лауреата Нобелевской премии среди советских химиков, и написал немало учебников для Высшей школы (в основном издательства Московского университета и издательства «Наука»), часть – со специалистом в области химической кинетики, деканом Химического факультета МГУ чл. – корр. АН СССР И.В. Березиным. За разработку кинетики химических и биологических реакций тот же автор стал лауреатом Государственной премии СССР по науке и технике (1984), а до того – лауреатом премии Ленинского комсомола (1978). Так что концептуально разработать количественную основу ДНК-генеалогии сделать было несложно, но надо было ее верифицировать на тысячах примеров, чтобы она была надежной. Это было сделано с использованием 3160 гаплотипов (2489 из них – в 67-маркерном формате, из 55 ДНК-генеалогических линий из 8 основных гаплогрупп). Эта работа и была опубликована в Advances in Anthropology в 2011 году[29], во втором выпуске журнала после его основания в том же году.
В последующем в том же журнале были опубликованы статьи с детальным описанием гаплогруппы R1a в мире, гаплогруппы R1a в Европе (с описанием 38 ветвей гаплогруппы R1a), гаплогруппы R1b, ДНК-генеалогии и лингвистики, ДНК-генеалогии и древней керамики от трипольской культуры до культур Юго-Восточной Азии и обеих Америк, а также четыре статьи об Африке и происхождении человека с точки зрения ДНК-генеалогии (две в 2012 г, и две – в 2014 г, одна из них редакторская). Одна статья из этих четырех[30] установила своеобразный рекорд популярности у читателей журнала – по состоянию на 25 октября 2015 года она была просмотрена более 90 тысяч раз, и «скачана» более 20 тысяч раз (http://www.scirp.org/journal/HottestPaper.aspx?Journalro=737).
Описанное выше – это крупный успех нового направления, отражение его признания. Стало ясно, что ДНК-генеалогия фактически является молекулярной историей (см. выше ответы на Вопросы 3 и 4).
Глава 2
Основные понятия и главные положения ДНК-генеалогии
Вопрос 16: Как в Y-хромосоме образуются мутации?
Время от времени при копировании Y-хромосомы в копии проскакивают ошибки. Фермент под названием ДНК-полимераза (на самом деле вместе с целым набором молекулярных инструментов копировальной машины) или просто ошибается и делает некоторые участки ДНК короче или длиннее, удлиняя или сокращая «тандемные повторы» нуклеотидов, или «ремонтирует» повреждённые участки и в ходе «ремонта» удаляет повреждение (эта операция называется «делеция»), или «вшивает» новый нуклеотид (называется «вставка», или «инсерт»). То, что удалось отремонтировать, в мутации не попадает, как будто мутации и не было. Что не удалось – передается сыну при рождении. Если родилась дочь, то Y-хромосома не передается. Если есть только дочери, или детей нет – Y-хромосома терминируется, вместе с мужской наследственной линией.
Мутации, которые интересуют ДНК-генеалогию, бывают двух типов – или изменение числа повторов, тандемов (STR), что описано выше, или «точечные», одно- или несколько-нуклеотидные (SNP), снипы.
Оказалось, что многие повторы нуклеотидов находятся у разных людей в одних и тех же участках Y-хромосомы. Эти участки уже специалистами пронумерованы, классифицированы, сведены в списки. Они получили название «маркеры». Набор маркеров, точнее, повторов в них, называется «гаплотип». Уже известных маркеров – больше тысячи. Они, как правило, одни и те же у всех людей на Земле и отличаются друг от друга только числом повторов, что вызвано теми самыми ошибками ферментов (и других элементов биологического копирования) при копировании из поколения в поколение.
Примеры повторов приведены в ответе на вопрос 14. Ясно, что «мутации», которые приводят к изменению числа тандемных повторов в Y-хромосоме, не есть те «мутации», о которых все имеют представление как о «поломках в генах», например, под воздействием радиации. Мутации, которые рассматривают в гаплотипах, происходят не в генах и не являются «поломками». Это, как объяснено выше – ошибки при копировании ДНК. Они имеют весьма сложную архитектуру, и радиацией не вызываются. Это – не поломка, а координированная перестройка.
Вопрос 17: Что такое «общий предок» выборки носителей гаплотипов, и как мутации могут помочь определить, когда он жил?
Любая группа мужчин, относящихся к одной и той же гаплогруппе и к одному и то же субкладу, а также к любой выборке гаплотипов, образующей ветвь в пределах одного субклада, являются в каждом случае родственниками, и происходят от одного общего предка. Можно привести и обратное определение – если мутация, которая осуществилась в Y-хромосоме, унаследовалась в ДНК потомков, то все эти потомки происходят от одного общего предка, у которого и произошла данная мутация.
В принципе, в любой гаплогруппе и любом субкладе имеется множество групп людей, которые происходят от их общего предка. Например, у двух родных братьев есть общий предок – их отец. Если к ним добавить дядю, то общий предок всех троих – отец дяди и дедушка братьев.
Чем глубже во времени общий предок группы людей, или популяции, в терминах ДНК-генеалогии, тем больше у него потомков. Все потомки наследуют гаплотип их общего предка, но чем дальше во времени отстоит общий предок, тем больше мутаций накапливается в гаплотипах потомков. Приведем простой пример. Допустим, общий предок жил 2500 лет назад, то есть 100 условных поколений назад (в ДНК-генеалогии за условное поколение принимается 25 лет, и константы скоростей мутации вычисляются в расчете на условное поколение протяженностью 25 лет). Если выявить сто потомков этого общего предка, в их 12-маркерных гаплотипах обнаружится примерно 200 мутаций, причем у 14 человек гаплотипы будут одинаковыми, и все 200 мутаций будут неупорядоченно распределены, в разной степени, между оставшимися 86 потомками.
Вот эти одинаковые 14 гаплотипов и представляют собой предковый гаплотип. За прошедшие 100 поколений – чисто статистически – в гаплотипах 86 потомков пройдут мутации, а в гаплотипах 14 потомков мутаций еще не будет. Вот таким образом определяют предковый, или базовый гаплотип.
Проверим представленные выше числа. 200 мутаций в 100 гаплотипах в 12-маркерном формате (константа скорости мутаций в 12-маркерных гаплотипах равна 0.02 мутаций на гаплотип за условное поколение) соответствуют величине 200/100/0.02 = 100 условных поколений, то есть примерно 2500 лет (здесь для простоты не вводится поправка на возвратные мутации, смысл которой будет пояснен в следующей главе). Это – линейный метод расчета времени до общего предка популяции. В то же время согласно логарифмическому методу если из 100 гаплотипов в 12-маркерном формате остались неизменными 14, то это соответствует времени, прошедшему от общего предка популяции, равному [ln(100/14)]/0.02 = 98 условных поколений, то есть примерно тому же времени, рассчитанному с помощью линейного метода.
Из этого примера должно быть понятно, что чем более недавний общий предок, тем большее число идентичных друг другу гаплотипов (или их фрагментов) наблюдается в выборке, и тем надежнее выявляется предковый гаплотип. Если общий предок жил очень давно, то одинаковых гаплотипов в выборке может остаться всего два-три – например, три гаплотипа из сотни 12-маркерных гаплотипов останутся при [ln(100/з)]/0.02 = 175 условных поколений до общего предка, то есть общий предок жил 4375 лет назад (на самом деле больше при учете возвратных мутаций). Когда в серии гаплотипов наблюдаются всего два-три одинаковых гаплотипа, они могут не обязательно быть предковыми, а оказаться одинаковыми случайно, в результате игры неупорядоченных мутаций. Поэтому понятие «предковый» гаплотип лучше не употреблять, за исключением совсем уже явных случаев, и в ДНК-генеалогии вместо «предковый» гаплотип обычно употребляют «базовый».
Итак, общий предок популяции – это предполагаемый носитель «базового», или «предкового» гаплотипа. Время жизни общего предка популяции вычисляется по совокупности гаплотипов его потомков, принимая во внимание число мутаций, накопившееся в популяции, нормированное на гаплотип или на маркер, и константу скорости мутации, или частоту мутации в расчёте на поколение. Расчёты времён жизни общих предков популяций позволяют делать предположения о времени и направлениях миграций популяций в древности, о передвижениях популяций. Поскольку, передвигаясь, популяции оставляли следы в виде материальных признаков, это позволяет проводить интерпретацию археологических данных в отношении носителей этих признаков, их принадлежности к определённым родам и племенам, связанных родственными взаимоотношениями. Аналогично, популяции, передвигаясь, приводили к перемещению языков, диалектов, и определение общих предков и времён их жизни даёт возможность прослеживать динамику языков в отношении соответствующих родов и племён, связанных родственными взаимоотношениями.
Вопрос 18: Можно ли привести примеры предковых гаплотипов, датируемых десятками и сотнями тысяч лет назад? Можно ли выявить гаплотип общего предка всех мужчин на Земле?
«Общий предок всех людей» – понятие скорее концептуальное, чем буквальное, абсолютное. Чем больше накапливается данных, тем понятнее становится, что «общий предок всех людей» теряется среди переплетений ДНК-генеалогических линий, уходящих на сотни тысяч лет вглубь. Поэтому введенное некоторое время назад понятие «хромосомный Адам» не является научным и никогда не было доказано. Интуитивно, такой общий предок возможен, но он, скорее всего, будет «плавающей мишенью».
Понятие «одного предка» можно рассматривать на разных уровнях сложности. Например, древнейшая африканская линия (гаплогруппа А00) имеет общего предка с гаплогруппой R1a примерно 210 тысяч лет назад, то есть оттуда, из такой глубины тысячелетий расходятся гаплотипы современных носителей гаплогруппы A00 и этнических русских гаплогруппы R1a:
13 11 12 10 11 16 10 8 14 14 8 8 8 9 12 11 12 8 12 12 11 11 (А00)







