




Полная версия
Достающее звено. Книга 2. Люди
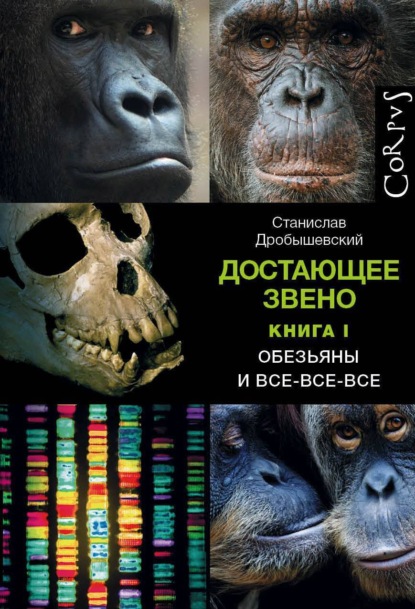

Систематика австралопитеков не так однозначна, как может показаться при чтении учебников. Южноафриканские антропологи, прежде всего Р. Кларк, давно аргументируют существование как минимум двух видов в Стеркфонтейне (например: Clarke, 2008). Один из них – “классический” Australopithecus africanus, тогда как второй – гораздо более крупный и массивный парантропоподобный, который, собственно, был предком массивных южноафриканских австралопитеков.
Уголок занудстваПредполагаемый парантропоподобный вид грацильных австралопитеков Южной Африки отличается целым комплексом черт: надбровье слабо выступает, вдавленность над ним минимальна, надглазничный край сравнительно тонкий; у самцов есть маленький сагиттальный гребень на задней части свода; затылок высокий и слабо выступающий назад; лицо очень высокое и длинное, вдавлено в поперечном направлении, скуловые кости резко выступают и развернуты вперед, при взгляде сбоку закрывают края носового отверстия; из-за этого лобный отросток верхней челюсти расширен вбок и ориентирован фронтально; межглазничное пространство широкое; точка “назион” (место соединения носовых костей с лобной) расположена выше лобно-верхнечелюстного шва, близко к “глабелле” (самой выступающей точке надбровья); нижняя челюсть большая и массивная; резцы и клыки увеличены относительно заклыковых зубов; премоляры и моляры большие, со вздутыми боками и бугорками, приближенными к центру зуба.
К этому виду относят черепа и челюсти Stw 252, Sts 71, Stw 505, Stw 183, Stw 498, Stw 384, Sts 1, MLD 2 и некоторые зубы.
При этом индивиды Sts 19, Sts 51 и Sts 52a, Stw 183, Stw 252, Stw 255 каждый в отдельности обладают специфическими чертами, иногда уникальными, иногда сближающими своих носителей с парантропами или “ранними Homo”. Одни исследователи относят это на счет индивидуальной изменчивости, другие – эволюционной переходности. Возможно, совершенствование методов датирования позволит прояснить ситуацию.
Впрочем, другие исследователи считают, что нет нужды разделять африканусов на два вида, а перечисленные выше различия объясняются половым диморфизмом, возрастной и индивидуальной изменчивостью или, возможно, хронологическими изменениями.
Заметим в скобках, что Р. Брум в 1936 году на основании разрушенного черепа Sts 60 из Стеркфонтейна описал вид Plesianthropus transvaalensis, а Р. Дарт в 1948 году по задней части мозговой коробки MLD 1 из Макапансгата – Australopithecus prometheus. Эти названия использовались некоторое время для обозначения грацильных австралопитеков из соответствующих местонахождений, но ныне обычно признаются полными синонимами Australopithecus africanus. Впрочем, некоторые исследователи считают название Australopithecus prometheus подходящим для обозначения того самого парантропоподобного вида, особенно если признать его бóльшую древность.
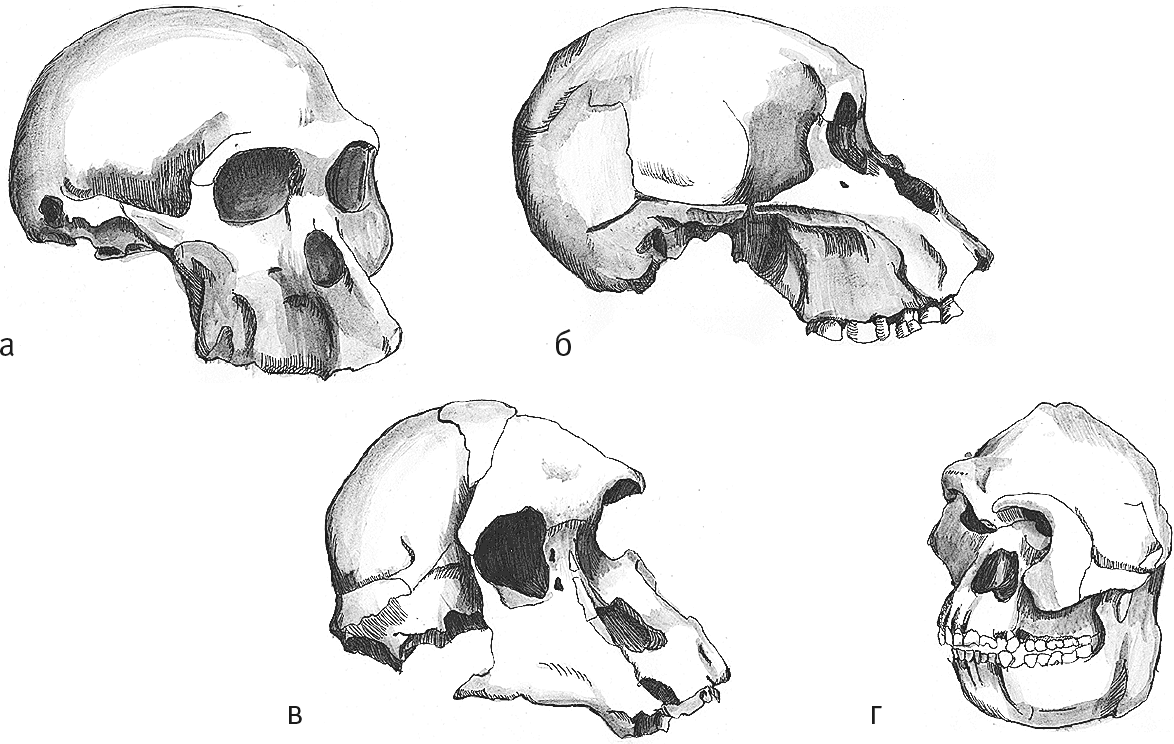
Рис. 10. Черепа Sts 5 (а), Sts 71 (б), Stw 505 (в), Stw 573 (г).
Другой важный вопрос – наличие Australopithecus africanus в Восточной Африке. К этому виду были отнесены грацильные австралопитеки из средних слоев Омо с датировками около 2,3 млн лет назад (например: Hunt et Vitzthum, 1986). Некоторые антропологи считают, что к африканусам можно причислить останки из местонахождения Южный Таквил (3,5 млн лет назад) и находки из Боури, описанные как A. garhi, в частности BOU-VP-12/130 (2,5 млн лет назад). С одной стороны, окаменелости из этих местонахождений очень фрагментарны, с другой – имеют отличия от “классических” афаренсисов из Лаэтоли и Хадара, с третьей – похожи на южноафриканских грацильных, но все же не совсем идентичны им. Учитывая, что грацильные австралопитеки Южной Африки в среднем моложе восточноафриканских, можно расценивать их различия как обусловленные временем, тогда находки из Омо и Боури логично вписываются в концепцию “хронологического A. africanus”. Если же считать, что грацильные австралопитеки Южной и Восточной Африки возникли независимо друг от друга, то надо допустить миграции африканусов с юга до Эфиопии около 2,5 млн лет назад, если не раньше.
Большинство современных антропологов считают, что A. africanus не участвовал в сложении нашего вида, но так думали не все и не всегда. В первой половине XX века, до открытий в Восточной Африке, африканусы были единственными известными обезьянолюдьми, и неудивительно, что многие ученые старательно выискивали в них человеческие черты. Так, черные прослои и пятна на костях из Макапансгата были приняты за следы древнейших очагов и обугленности. После выяснилось, что это окислы марганца. Р. Дарт собрал впечатляющую коллекцию костей, рогов и зубов из южноафриканских пещер и описал остеодонтокератическую – костно-зубо-роговую – культуру (Dart, 1957). Сломы на концах костей и рогов он принял за результаты использования, а разрушения на черепах павианов – за следы ударов костями (причем якобы повреждено аж 80 % найденных черепов, а расположение проломов позволило утверждать о праворукости охотников), а также считал, что двойные вмятины на некоторых из этих черепов соответствуют мыщелкам длинных костей копытных, найденных рядом.
Надо отдать Р. Дарту должное, он не был голословен: среди богатых этнографических коллекций, собранных среди бушменов, он обнаружил удивительно точные параллели остеодонтокератическим обломкам. Некоторые изделия, кажется, делал один человек, только вот на бушменских есть орнамент, а на австралопитечьих – нет. Однако исследование некоторых образцов из “остеодонтокератической” коллекции новейшими методами показало, что характерные повреждения их концов – это погрызы гиен, а не рабочая изношенность. Так что, видимо, “культура” австралопитеков на самом деле является объедками падальщиков; впрочем, проверялись далеко не все собранные Р. Дартом кости, так что надежда на реабилитацию южноафриканских австралопитеков еще остается.
Светлым лучиком в беспробудной тьме, окружающей интеллект австралопитеков, является одна уникальная находка: в Макапансгате в брекчии с датировкой 2,5–3 млн лет найден круглый камень красного цвета – размером с кулак без следов обработки. Самое интересное, что ближайшее местонахождение такой породы расположено за 32 км от пещеры. Ясно, что камень не сам прополз эту дистанцию. А ведь это целый дневной переход – немалое расстояние, тем более что нести камень надо было все время в руке, корзин и мешков-то еще никто не изобрел! Более того, естественные выщерблины на камне напоминают глаза и рот, образуя чудную рожицу. Как знать, может, некий первобытный эстет, гуляя по родной степи, усмотрел в этом булыжнике свое отражение и не поленился притащить его домой? Камень из Макапансгата – древнейшее “прикольно” или “хочется чего-то этакого, чтоб душа развернулась, а потом обратно завернулась”. А может, это самый первый смайлик?
Мозговитый Беби из Таунга – несостоявшийся прогресс?Чем больше антропологи смотрят на старые находки, тем больше интересного видят. Ныне взгляд обратился на дартовского Беби из Таунга – классический череп детеныша австралопитека, первый в великолепном ряду последующих открытий. Кажется, за предыдущие без малого 90 лет про Таунга выяснили все что можно, например, что его заклевал злой орел. И вот теперь международная (какая же еще в век глобализации?) группа исследователей заострила свое внимание на наличии следов метопического шва на эндокране – естественном слепке мозга – Таунга. Метопический шов – это шов между двумя половинками лобной кости. То есть лобная кость у ребенка вообще-то изначально парная, но правая и левая части обычно срастаются примерно к двум годам. Однако же следы шва, а то и он весь регулярно остаются незаращенными. Этот факт всегда обращал на себя внимание антропологов, но в палеоантропологическом аспекте им занимались мало, обычно просто констатируя случаи сохранения шва на взрослых черепах.
В нынешнем же исследовании ученые сравнили частоту незаращения шва у шимпанзе, ископаемых гоминид и современных людей (Falk et al., 2012). Выяснилось, что у обезьян метопический шов закрывается почти у всех детенышей к моменту прорезывания второго молочного моляра, а ко времени прорезывания третьего постоянного у всех поголовно особей исчезают даже его следы. У современных же людей, хотя между первым и вторым молочными молярами частота наличия шва резко падает, частенько шов или его следы остаются на всю жизнь.
У ископаемых гоминид картина тоже весьма показательна. Среди массивных австралопитеков – южно- и восточноафриканских – не известно ни одного случая сохранения хотя бы следов шва. А вот среди грацильных австралопитеков, “ранних Homo”, эректусов и неандертальцев частота шва или его следов весьма высока, видимо, не меньше, чем среди современных людей.
Авторы статьи предположили три причины такой ситуации. Во-первых, шов мог сохраняться для облегчения родов большеголовых детенышей прямоходящими самками. Правда, остается неясным, зачем тогда шву быть открытым многие годы после этого радостного события? И почему тогда парантропы остались в стороне от прогресса? Мозгов у них было примерно столько же, что и у грацильных австралопитеков, а челюсти – крупнее. Во-вторых, долгое сохранение шва после родов могло способствовать росту мозга у детеныша. Наконец, в-третьих, оно может отражать прогрессивную перестройку новой коры головного мозга, в особенности лобной доли – самой продвинутой, отвечающей за мышление как таковое. Очевидно, все эти причины взаимосвязаны и, скорее всего, действовали одновременно. В этом свете логично, что шимпанзе и парантропы рано обретают “твердолобость” – жевательные мышцы для них важнее интеллекта.
Красиво, хотя моя ехидная память подсказывает одно маленькое но. Помнится, в 1995 г. одна из авторов нынешней статьи – Дин Фальк, известная своими исследованиями ископаемых эндокранов, в составе авторитетного авторского коллектива доказывала, что Беби из Таунга с наибольшей вероятностью – детеныш не африканского австралопитека, а парантропа (Falk et al., 1995). Уникальная закавыка систематики, если учесть, что Беби – голотип Australopithecus africanus! Впрочем, может, среди парантропов были свои прогрессивные представители. С другой стороны, он же погиб ребенком, явно не оставив своих продвинутых генов в потомстве; крючковатые когти орла пресекли прогресс парантропов… Родители-то были еще дремучие, не отмахались от грозной птицы, не уберегли драгоценное чадо! Приятно сознавать, что наши предки, обогащенные позднесрастающимся метопическим швом, оказались смышленее и прогрессивнее. Не то эту статью писал бы этакий клацающий челюстями “щелкунчик”-постпарантроп. Однако повторим великую истину: палеонтология не терпит сослагательного наклонения.
Кстати, об ужасах систематики…Как было упомянуто выше, Беби из Таунга запросто может быть детенышем массивного австралопитека (Falk et al., 1995). В пользу этого говорят три основных признака, типичных именно для парантропов: во-первых, увеличенный затылочно-краевой синус – сосуд на задней стороне мозга для оттока венозной крови; во-вторых, наднебное расширение верхнечелюстных синусов – полостей в теле верхней челюсти; в-третьих, сравнительная площадь бугорков на первом нижнем моляре. По всем этим показателям Таунг оказывается парантропом. Но можно ли тогда вообще называть его этим словом? Ведь Таунг – голотип вида Australopithecus africanus, значит, это название по правилу приоритета должно использоваться для обозначения южноафриканских массивных австралопитеков. Тогда южноафриканские грацильные будут носить имя Plesianthropus transvaalensis, восточноафриканские грацильные – Plesianthropus afarensis, нынешние анаменсисы должны бы именоваться Plesianthropus africanus (учитывая название Meganthropus africanus, данное Г. Вейнертом в 1950 г. челюсти из Гаруси), а восточноафриканские массивные – Australopithecus boisei. И это без учета мнений, согласно которым восточноафриканские грацильные достойны собственного родового названия Praeanthropus, а восточноафриканские массивные – Zinjanthropus.
Вот к чему приводят случайности! Если бы Р. Дарту первым попался череп взрослого австралопитека, этой ужасной путаницы удалось бы избежать. А теперь систематики вынуждены иногда просыпаться в холодном поту и с липкими руками – когда им снится кошмар, что кто-то особо въедливый и принципиальный поставил-таки вопрос ребром о пересмотре названий. Избежать этого ужаса можно, доказав, что все же Таунг – грацильный австралопитек, или переопределив голотип. Впрочем, антропологи осознают, какие названия и по отношению к кому они используют, друг друга они понимают, а потому живут без лишних формальностей.
На исходе века, в 1999 году, научный мир узнал о новом виде австралопитеков…
Австралопитек гари Australopithecus garhi – крайне любопытное существо, фрагменты черепа и конечностей которого найдены в Эфиопии в местонахождениях Боури, Гамеда, Матабайету, а также, возможно, Ндоланья и Кооби-Фора и имеют возраст 2,5 млн лет (Asfaw et al., 1999). Это – самый поздний из грацильных австралопитеков. Здорово, что в Боури обнаружен фрагментарный скелет BOU-VP-12/1, отлично описанный (DeGusta, 2004), только отчего-то непропиаренный и потому малопопулярный.
Череп BOU-VP-12/130 своеобразен: лобная кость с достаточно мощным надбровьем, а височные линии, видимо, еще на лобной кости сходились в сагиттальный гребень, который заканчивался уже на теменной, – такой вариант резко отличается от версии A. afarensis и A. africanus, у которых гребень был смещен далеко назад. Уникальна верхняя челюсть: альвеолярный отросток выпуклый как поперечно, так и продольно, крайне сильно выдается вперед, отчего его передний конец аж поднимается наверх, а резцы оказываются торчащими вперед. Такого прогнатного гоминида планета не видела ни до, ни после. Форма альвеолярной дуги – как верхней, так и нижней – крайне архаичная, U-образная, больше всего похожая на типичную для A. anamensis – гораздо более древнего вида. Другой особенностью A. garhi являются очень большие зубы – как задние, так и передние. Моляры этого вида сопоставимы со значениями не самых крупных парантропов, но вот огромные резцы ставят A. garhi явно особняком.
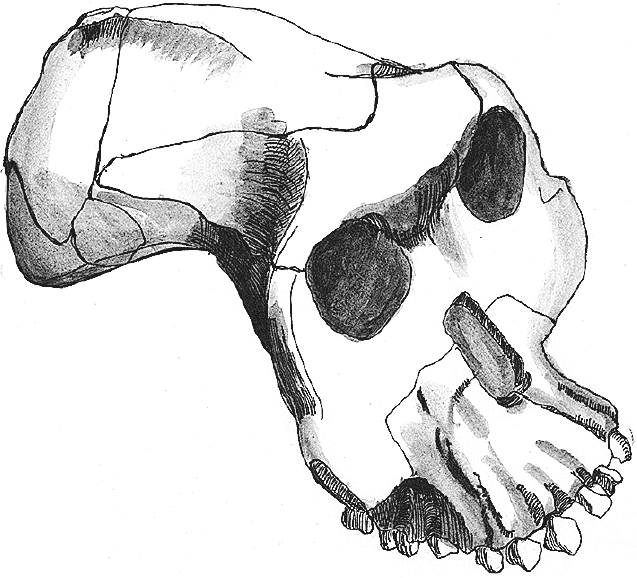
Рис. 11. Череп Australopithecus garhi.
Крайне интересно, что посткраниальные кости A. garhi больше похожи на таковые “ранних Homo”, чем A. afarensis. Это довольно неожиданно, коли уж гари не был нашим предком. Закономерно же то, что по строению скелета A. garhi родственнее восточноафриканским гоминидам, чем южноафриканским.
Рост A. garhi был стандартнейшим для австралопитеков – от метра до полутора. А вот пропорции конечностей BOU-VP-12/1 уникальны, такие не встречены больше ни у каких гоминид: руки были, видимо, резко удлинены относительно ног, причем в основном за счет увеличенных предплечий.
Получается, что гари был каким-то очень странным австралопитеком – последним грацильным, но очень уж крупнозубым, специализированным по черепу, но продвинутым по скелету. Он явно не входил в число наших предков, иначе пришлось бы допустить быстрое увеличение зубов и челюстей, а после такое же быстрое уменьшение, да и специализации его слишком своеобразны.
Как уже говорилось выше, австралопитеки гари с большой вероятностью изготавливали каменные чопперы и умели разделывать ими антилоп. А ведь к этому времени уже появились “ранние Homo” – наши прямые предки. Выходит, в Восточной Африке существовали как минимум две независимые, хотя и родственные группы, находившиеся на пути очеловечивания. Судя по тому, что Читатель этой книги явно не потомок австралопитеков гари, победили Homo, хотя мы и не знаем почему. Плюс к этому появившиеся тогда же массивные австралопитеки несколько позже тоже, видимо, стали изготавливать каменные орудия, причем восточноафриканские делали это независимо от южноафриканских.
Кстати, о массивных австралопитеках…
Глава 3
Щелкунчики, побежденные мышами: массивные австралопитеки
Около 2,5 млн лет назад начались новые глобальные изменения климата. Ландшафты Африки стали еще более открытыми, широко распространились саванны. Вместе с ландшафтами изменилась и фауна. Эти преобразования коснулись и австралопитеков. Появились сразу две группы – “ранние Homo” и массивные австралопитеки, или парантропы.
Парантропы не были нашими предками, но их изучение интересно, так как дает возможность взглянуть на историческую альтернативу, параллельную нам эволюционную ветвь. Даже само название говорит о том же: “парантроп” буквально значит “околочеловек”. Это уникальные существа, не имеющие никаких аналогов в современной фауне.
Предками восточноафриканских массивных австралопитеков были, по-видимому, Australopithecus afarensis. В облике первых парантропов очевидным образом прослеживается план строения афаренсисов. Фрагмент свода черепа KNM-ER 2602 с датировкой 3,3 млн лет назад описывался и как Australopithecus afarensis, и как Paranthropus aethiopicus, так что он может считаться достающим звеном между этими видами (впрочем, в нем же подозревали и Kenyanthropus platyops). Фрагменты верхних челюстей Omo L 55-s-33 и EP 1500/1 намного моложе – 2,6–2,7 млн лет назад, они уже явно принадлежали массивным австралопитекам. А челюсть Omo L 18-1967-18 из еще более позднего уровня (2,5 млн лет назад) послужила голотипом для описания рода и вида Paraustralopithecus aethiopicus (Arambourg et Coppens, 1968). В дальнейшем ранг древнейшего вида массивных австралопитеков был понижен, и ныне он называется…
Парантроп эфиопский Paranthropus aethiopicus известен из нескольких восточноафриканских местонахождений – Кооби-Фора, Ндоланья, Омо, Локалеи – с датировками от 2,7 до 2,3 млн лет назад. Особенно знаменит “Черный череп” KNM-WT 17000, уникальный своей сохранностью. Он очень похож на череп афарского австралопитека, но его челюсти гораздо крупнее, а моляры больше, хотя и недотягивают до значений, типичных для поздних парантропов. От последних “Черный череп” отличается сильнейшим прогнатизмом: выступанием челюстей вперед он превосходит даже горилл! Тяжелые челюсти приводились в действие мощными жевательными мышцами, крепившимися на сагиттальном гребне. Крупная морда уравновешивалась шейными мышцами, тянувшими за выйный гребень. Лицо эфиопского парантропа широкое, резко уплощенное, даже вдавленное, с огромными скуловыми дугами, не достигавшими, впрочем, степени развития Paranthropus boisei. Объем мозга KNM-WT 17000 – 410 см³, а форма мозга типична для парантропов; в частности, височная доля выступает очень слабо. В отличие от позднейших парантропов, Paranthropus aethiopicus имел крупные клыки и не слишком маленькие резцы.
Про этот вид известно не так уж много. Другие находки намного фрагментарнее. Кстати, выдвигалось предположение, что череп BOU-VP-12/130 принадлежал самке P. aethiopicus, но его специфика все же слишком велика, а сходство объясняется, видимо, происхождением от одного предка – афарского австралопитека.
Сами же парантропы быстро эволюционировали и уже спустя несколько сотен тысяч лет превратились в новый вид…
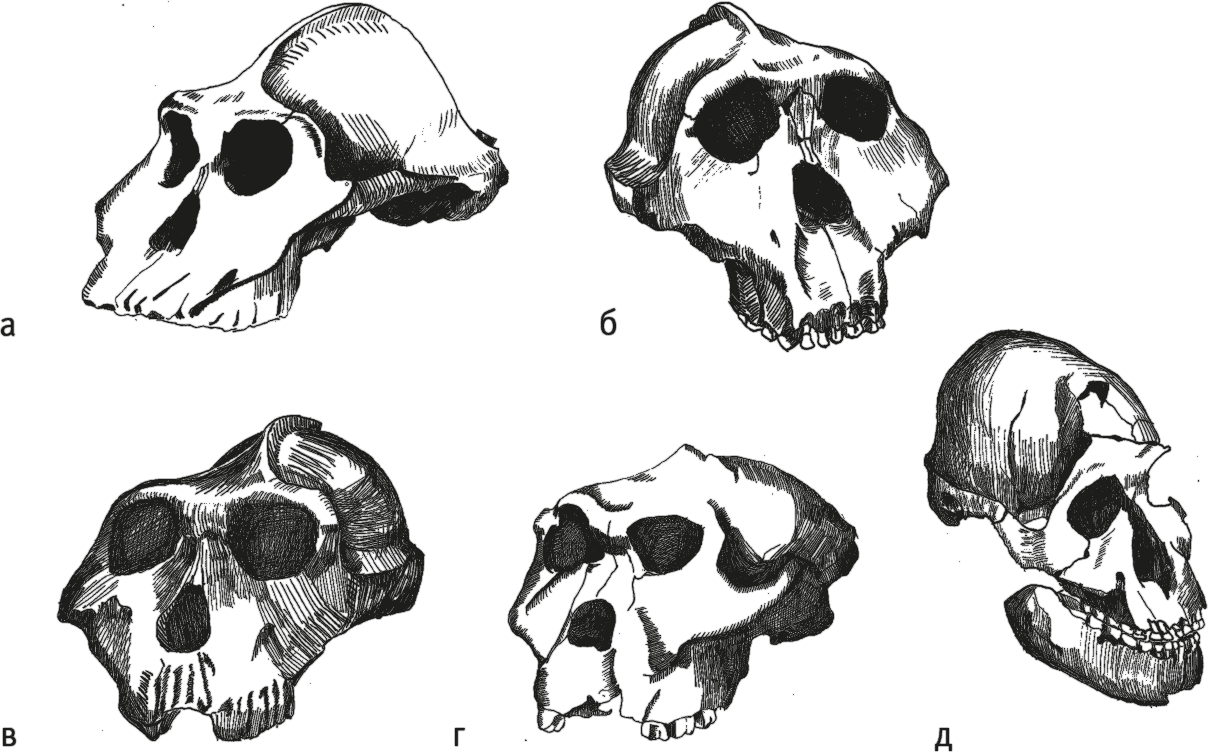
Рис. 12. Череп Paranthropus aethiopicus KNM-WT 17000 (а), черепа Paranthropus boisei OH 5 (б) и KNM-ER 406 (в), черепа Paranthropus robustus SK 48 (г) и DNH 7 (д).
Парантроп Бойса Paranthropus boisei обитал в Восточной Африке, на территории нынешних Танзании, Кении и Эфиопии в интервале примерно от 2,5 до 1,1 млн лет назад. Самые ранние P. boisei трудноотличимы от P. aethiopicus, так что многие находки имеют двойное определение. Около 2–1,5 млн лет назад это был едва ли не самый массовый вид австралопитеков, о чем говорит и обилие местонахождений – Омо, Каитио, Кангатукесео III, Нату, Чесованжа, Консо, Кооби-Фора, Олдувай, Малема, Пенинж. Сами находки тоже радуют, ведь в нашем распоряжении целый ряд почти целых черепов – OH 5, KGA10–525, KNM-ER 406, KNM-ER 732, KNM-ER 13750, KNM-ER 23000, Omo L 323-1976-896, KNM-WT 17400, KNM – CH 1, не говоря об отдельных челюстях, зубах и посткраниальных костях (Wood et Constantino, 2007).
Массивные австралопитеки неспроста получили свое название. И неспроста первый описанный восточноафриканский череп – OH 5 – получил прозвище Щелкунчик (Leakey, 1959). Черепа бойсовских парантропов очень мощные, крупные, с развитыми гребнями для жевательной и шейной мускулатуры – сагиттальным и затылочным (даже у самок и подростков!), очень узким и плоским лбом, которого как бы даже и нет, довольно слабым надбровьем (выступающим, впрочем, сильнее, чем у грацильных австралопитеков), а также широко развернутыми толстыми скуловыми дугами. Дуги эти винтообразно изогнуты, за счет чего лишний раз укреплены. При взгляде сверху череп P. boisei ни с чем невозможно спутать, он похож на экзотический древнегреческий кувшин: надбровье выполняет роль венчика, экстремально выраженное позадиглазничное сужение – горлышка, скуловые дуги – ручек, мозговая коробка – тулова сосуда, а поперечный затылочный гребень – донца. От своих предков бойсовцы унаследовали уплощенность лица и развернутость скул вперед. Судя по всему, существовали высоко- и низколицые варианты, хотя пока неясно, привязаны ли они ко времени или географии. Челюсти парантропов просто огромные, но при этом ортогнатные, то есть не выступают вперед – так нагрузка на челюсти получается большей. Самая главная особенность парантропов – специализированная зубная система: резцы и клыки у них были очень маленькими, а вот премоляры и моляры – чудовищных размеров, причем морфологически премоляры очень похожи на моляры.
Очевидно, бойсовцы были специализированы по типу питания. Огромные зубы и челюсти явно были даны им неспроста. Как говаривал Винни-Пух: такой пастью, да медку бы! Но нет, изотопные анализы и микростертость эмали свидетельствуют о какой-то жесткой травянистой прибрежной растительности типа осоки. Надо сказать, до поры до времени такая экологическая ниша себя отлично оправдывала, неспроста останки парантропов встречаются в отложениях в разы чаще, чем “ранних Homo” (есть, правда, вероятность, что такое распределение обусловлено не численностью, а образом жизни и тафономией: парантропы могли жить на берегах и чаще попадать в воду после смерти, а “ранние Homo” – на удалении от водоемов и, соответственно, будущих геологических слоев). Впрочем, специализация специализацией, но экологическая пластичность приматов проявлялась и в парантропах: в местонахождении Пенинж, где найдена челюсть P. boisei, условия были самыми сухими из всех известных в Африке, а ландшафт представлял открытую травяную саванну.
Да и с интеллектом у них могло быть не все так печально, как может представиться из образа меланхоличного осокопожирателя. Мозг P. boisei был крупнее, чем у грацильных австралопитеков, – в среднем около 490 см³ с размахом от 390 до 545 см³. Его строение, правда, очень примитивное – с маленькой и резко суженной лобной долей, слабо выступающей височной, но все же чуть продвинутее современных шимпанзе. Такой уровень, весьма вероятно, позволял им изготовлять орудия труда, в том числе каменные. Другое дело, что при их образе жизни эти орудия были, надо думать, не очень-то и нужны: травы мало не бывает, рвать ее можно и руками, а от хищников удобно прятаться на деревьях.
Судя по посткраниальным костям, строением тела парантропы не слишком отличались от своих грацильных предков, с поправкой на массивность и размеры. P. boisei были крупными по австралопитековым меркам: до 1,24–1,56 м ростом и 34–49, возможно, даже до 70 кг весом. Половой диморфизм парантропов был выражен довольно сильно; по размеру самки были в полтора раза меньше самцов, а на их черепах височные линии могли не сливаться в сагиттальный гребень.
Древесный парантроп?Группа массивных австралопитеков – парантропов – известна уже давно, почти сто лет. В Южной и Восточной Африке за это время найдены едва ли не тысячи зубов и многие сотни обломков черепов и прочих костей от нескольких сотен индивидов. Но вот что удивительно: до сих пор ни одного даже фрагментарного скелета не было найдено! Это может показаться загадочным, тем более что парантропы были даже более многочисленными, чем, например, “ранние Homo”. Однако факт остается фактом – скелетов нет. Объясняется сия странность главным образом обстоятельствами смерти парантропов. Слишком часто они попадали на ужин местным хищникам. А ведь только в дешевых комиксах, на карикатурах и неудачных реконструкциях рядом с логовом ужасных хищников-людоедов лежат аккуратные скелетики. В действительности львы и леопарды разрывают добычу на мелкие клочки, а гиены и шакалы обычно не оставляют после себя ни одной целой кости. Одна и есть надежда у палеонтологов – неаккуратные крокодилы, щедро раскидывающие куски тела жертвы, да еще в воде, где их не могут достать падальщики и где идеальны условия захоронения. К сожалению, древние гоминиды нечасто лезли плавать в пруды с родичами динозавров.







