


Полная версия
СТРУКТУРНАЯ ОРГАНИЗАЦИЯ ХРОМОСОМ И ДИФФЕРЕНЦИРОВКА КЛЕТОК КРОВИ. STRUCTURAL ORGANIZATION OF CHROMOSOMES AND BLOOD CELL DIFFERENTIATION
Известно, что упаковка ДНК происходит специфически и с учетом активности генов (Храпунов С. Н., Драган А. И., Бердышев Г. Д., 1987). Следовательно, в популяции костного мозга неадекватность линейных параметров и различную организацию морфологических структур хромосом в разных типах наборов определяется направление дифференцировки и работа генов с учетом будущей специализации и образования лейкоцитов, эритроцитов и мегакариоцитов. Так проявляются индивидуальные функционально-генетические установки на дифференцировку разных типов специализированных клеток.
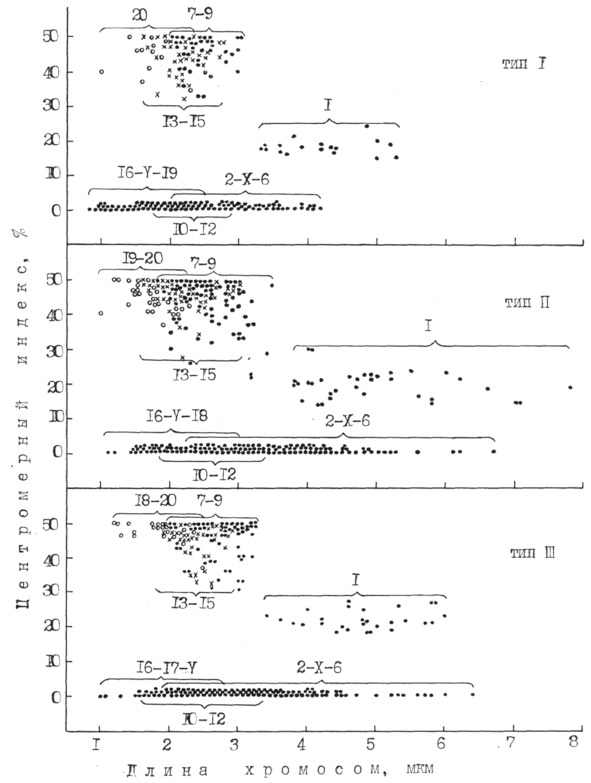
Таким образом, классификация хромосомных наборов из костного мозга с учетом их линейных параметров и морфоструктуры позволила выделить три типа хромосомных наборов, различающихся количеством акроцентрических и метацентрических хромосом.
Метафазная хромосома – это плотно упакованная нить ДНК, которая в стволовых клетках имеет одинаковый набор генов. При митозе набор генов в клетках не меняется, но в специализированных эритроцитах, лейкоцитах и мегакариоцитах изменяется соотношение неактивных и активных генов, кодирующих образование различных белков.
Эти изменения закладываются через упаковку ДНК на стадии интерфазы, когда определяется направление дифференцировки клеток в соответствии с потребностями организма и его физиологией, что затем сказывается на организации морфологической структуры метафазных хромосом в разных клетках.
Итак, в костном мозге происходит образование трех основных морфологических типов кроветворных клеток: лейкоцитов, эритроцитов и мегакариоцитов и направление к их дифференцировке осуществляется через образование трех типов хромосомных наборов.
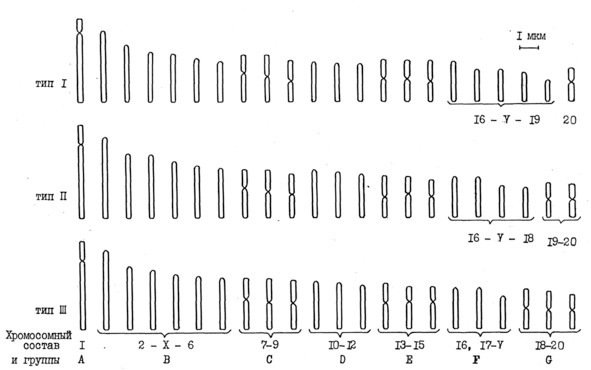
3.2. Сравнение структурной организации хромосомных наборов из культуры крови и костного мозга
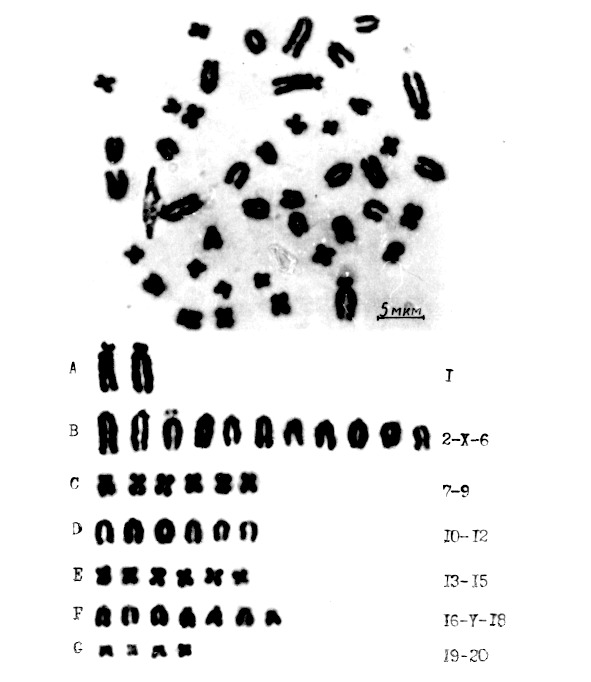
Для изучения структурной организации хромосом в лейкоцитах крови была поставлена культуры клеток крови от 4 самцов беспородных белых крыс. Визуально проанализировано 52 хромосомных набора, и все они имели структурную организацию по типу II (рис.5).
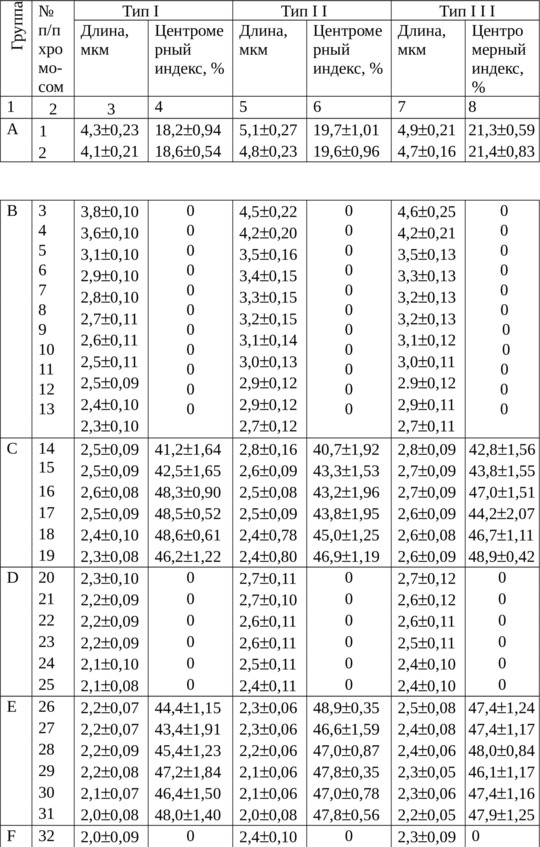
Для морфометрической идентификации использовали 16 метафазных пластинок. Оценку достоверности степени спирализации провели по средней суммарной длине хромосом в группах и критерию достоверности (табл. 3). Этот подход согласуется с мнением А. Р. Круминя (1989), который считает, что самым точным показателем степени спирализации может служить суммарная длина хромосомного набора, но такая работа трудоемка.
Рис. 5. Структурная организация хромосом в культуре клеток
периферической крови белой крысы самца
В культуре крови, как известно, обладают способностью к делению лимфоциты (Бочков Н. П., 1971; Олиници К. Д.,1982), и нами там выявлен лишь один тип II структурной организации хромосом. В костном мозге, где образуются три ростка кроветворения (Абрамов М. Г.,1979, 1985), нами выделены три основных типа структурных организаций хромосом.
При сравнении морфометрических показателей хромосом из крови и костного мозга не обнаружено различий по центромерному индексу, но линейные параметры хромосом из крови были меньше. Длина в среднем у самой большой и наименьшей хромосомы из культуры крови была 4,2±0,12 и 1,1±0,05 мкм, а в этом типе из костного мозга – 5,1±0,37 и 1,6±0,08 мкм с достоверностью разности между ними соответственно при Р> 0,95 и P> 0,999.
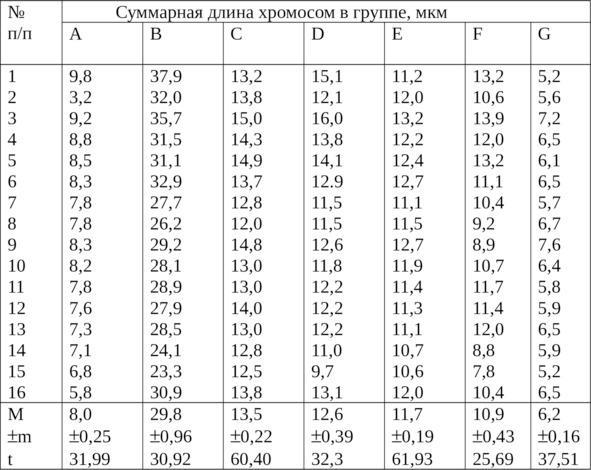
Суммарная длина хромосом в группах из клеток культуры крови представлена в таблице 3.
Наименьшую вариабельность по суммарной длине наблюдали у метацентрических хромосом групп С, Е и G, у которых был наибольший критерий достоверности (t): 60,4; 61,9 и 37,5.
Таблица 3
Суммарная длина хромосом в группах из культуры клеток крови
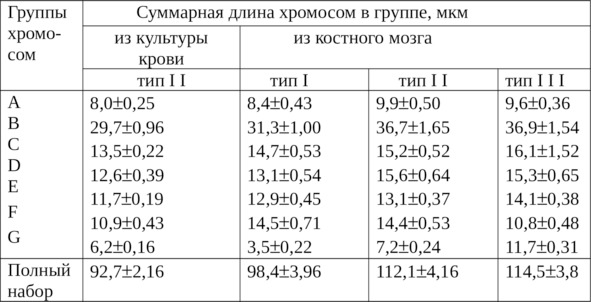
Сопоставление линейных параметров из культуры клеток и костного мозга показано в таблице 4. Если в костном мозге длина всех хромосом в наборе была наименьшей в типе I – 98,4±3,96 мкм и наибольшей в типе III – 114,5±3,78 мкм, то в клетках из культуры крови она составила лишь 92,7±2,16 мкм. Это было значительно меньше суммарной длины хромосом типа II из костного мозга, где равнялась 112,1±4,16 мкм.
Зафиксированные различия на стадии метафазы по структурной организации хромосом в наборах из культуры крови и костного мозга можно объяснить лишь различными условиями пролиферации.
Таблица 4
Сопоставление линейных параметров хромосом
из культуры крови и костного мозга
Д. Конки и соавторы (1989) отмечали, что размножение и дифференцировка клеток требуют различных условий.
Процессы дифференцировки родоначальных кроветворных клеток в костном мозге, как известно, осуществляются при влиянии различных гормонов и других уровней регуляции с учетом потребностей организма (Моничев А. Н., 1984; Гаврилов О. К., Фанштейн Ф. Э., Турбина Н. С., 1987). В культуре крови отсутствуют сложные уровни регуляции, имеющиеся в организме, поэтому там образуется один тип хромосомных наборов – тип II. В костном мозге, где происходит кроветворение и дифференцировка клеток в разных направлениях при одном и том же генном наборе, что и в культуре крови, образуются три типа хромосомных наборов.
На стволовые клетки в организме оказывают влияние различные процессы контактных и дистанционных биохимических воздействий с учетом внутренней среды организма и его потребностей. Следовательно, этот сложный механизм взаимодействия обуславливает в костном мозге формирование трех типов хромосомных наборов.
3. 3. Идентификация трех типов хромосомных наборов при дифференциальных окрасках
На равномерно окрашенных хромосомных наборах из костного мозга четко прослеживается различие по числу акроцентрических и метацентрических морфоструктур, но гомологичные пары достоверно идентифицировать можно лишь некоторые.
С целью более четкой идентификации пар аутосом и выделения половых хромосом мы провели G-окраску с применением протеолитического фермента трипсина. При этом мы не получали всегда идентичной G-окраски половых хромосом и аутосом как в трех типах хромосомных наборов, так и в пределах типа. Сегментная проработка структуры хромосом во многом зависела от дозы и времени действия колхицина и фермента (рис. 6).
На фотографиях одного и того же набора типа II показано, что при малом воздействии фермента (рис. 6а) имеется некоторая сегментация у первой субтелоцентрической пары и отдельных акроцентрических хромосом, у метацентрических хромосом сегменты почти не выражены.
Конец ознакомительного фрагмента.
Текст предоставлен ООО «Литрес».
Прочитайте эту книгу целиком, купив полную легальную версию на Литрес.
Безопасно оплатить книгу можно банковской картой Visa, MasterCard, Maestro, со счета мобильного телефона, с платежного терминала, в салоне МТС или Связной, через PayPal, WebMoney, Яндекс.Деньги, QIWI Кошелек, бонусными картами или другим удобным Вам способом.