


Полная версия
Collins New Naturalist Library
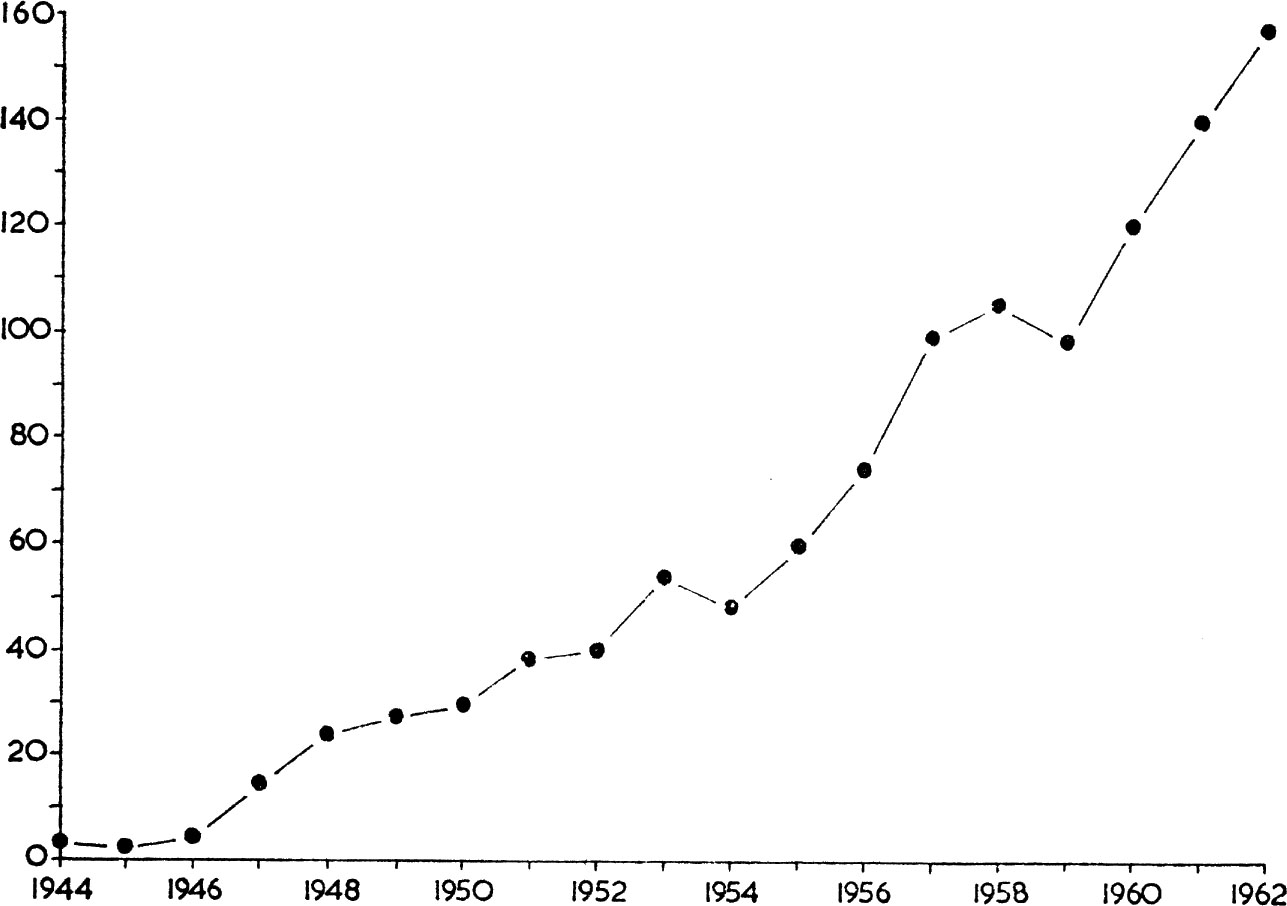
FIG. 6. Increase in number of pairs of little ringed plovers summering in Britain. (Data from Parrinder 1964).
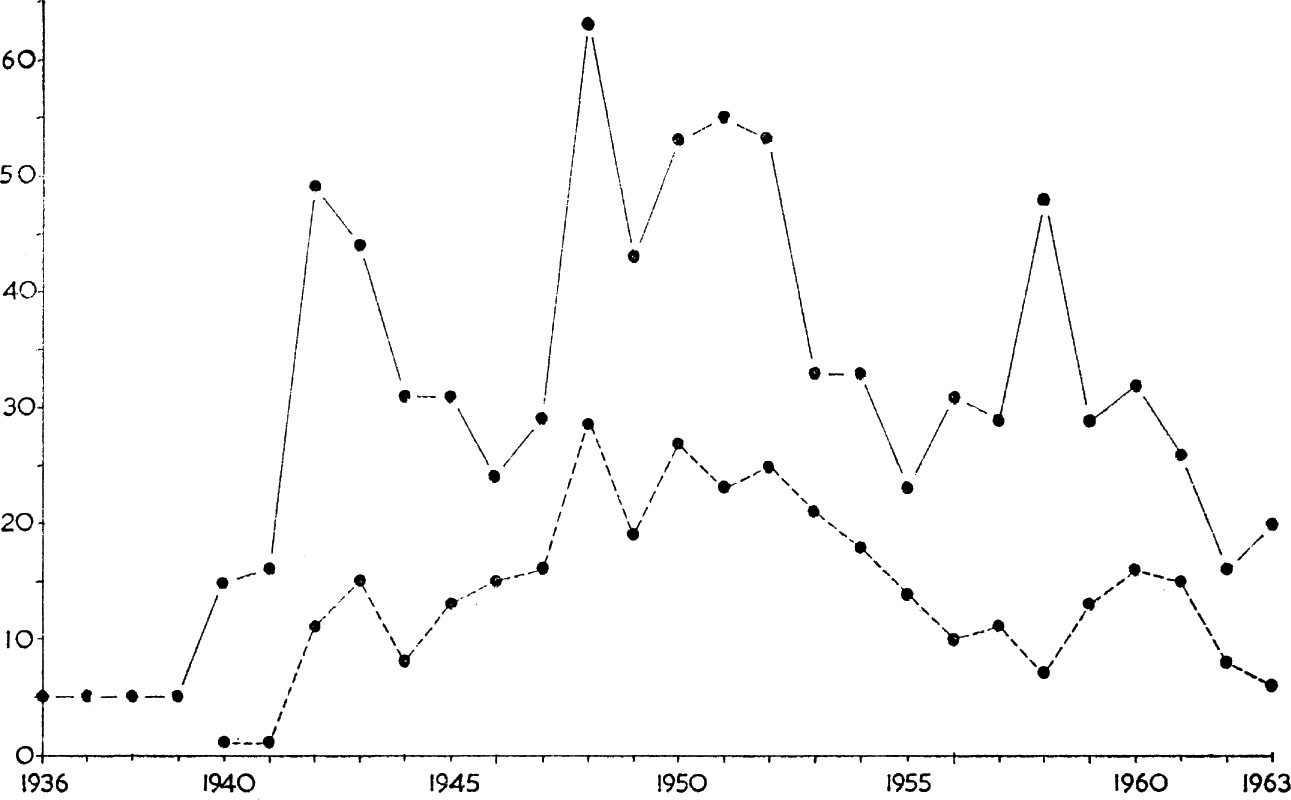
FIG. 7. Changing status of black redstart in Britain showing increase in number of territory holding males during the 1939–45 war with a decline following the final clear-up of war damaged sites after 1950. The solid line gives figures for the whole of Great Britain, while the dotted line is the contribution made by the City of London and Dover combined. (Data from Fitter 1965).
The black redstart was originally a bird of the warm montane regions of the southern Palaearctic, like the rock thrush and crag martin, but spread northwards after the last glaciation. It has adapted itself to a man-made environment, using buildings for nesting sites in lieu of cliff faces, and in parts of Germany it replaces the robin as the familiar follower of man. Its northward spread was still in progress across Germany during the last hundred years, and it only reached Jutland in the second half of the nineteenth century, and Scandinavia in the early 1940s. There is no record of it in Britain before 1819, though it became a regular passage migrant on the south and east coasts during the middle of the nineteenth century. Sporadic nesting attempts followed, in Durham in 1845, Sussex in 1909 and then a slow build-up from 1923 onwards. Three pairs nested at the Palace of Engineering in Wembley, Middlesex from 1926–41 but altogether less than half a dozen pairs were breeding in Britain up to the mid 1930s after which a big increase occurred, detailed in Fig. 7. The ability of the species to establish itself in this way has followed the sudden availability of nesting sites and feeding grounds, in the form of war-time bombed sites, particularly in London and Dover. Rebuilding and the post-war clean-up account for the subsequent decline in numbers.
There are also examples of bird numbers reduced through direct persecution, especially when the victim is fairly rare. The great crested grebe was certainly widely distributed in suitable places in Britain in the early nineteenth century, but by the middle years of the century a big demand arose for its breast feathers to make ‘grebe furs’ for a fashionable home market, and the slaughter began in 1857. By 1860, the species was reduced to 42 known pairs and was only saved by the sanctuary afforded by private estates, while further help came with the Bird Protection Acts of 1870–1880. Nevertheless, some increase was under way by 1880, before protection could have been very effective, and Harrison and Hollom (1932), who record these early changes, consider that human persecution came at the start of a period of long-term cyclical increase. By 1931, there were around 1,150 breeding pairs (with non-breeders, about 2,650 adults) in England and Wales and about another 80 pairs in Scotland. A sample census by Hollom (1959) showed that about the same number of adult grebes existed in Britain in the 1940s, but that an increase then began. When Prestt and Mills undertook a census in 1965 there were approximately 4,500 adults in Britain. This increase seems to have been favoured by man’s activities in creating numerous new reservoirs and gravel pits, just as the little ringed plover benefited. The 70% increase of the population in about twenty years can be compared with an increase of 84% in sand and gravel production between 1948 and 1957. That an increase followed the creation of new habitats also indicates that saturation had previously been attained and that the bird was regulated in the sense already discussed.
For a species to extend its range and take advantage of newly developed habitats it would be helpful for it to possess some kind of exploratory behaviour rather than rely on chance movements. It is becoming clear that immediately after breeding, many species, which normally migrate south, first indulge in northerly flights. The large-scale ringing of sand martins has shown that birds breeding at a colony in the south of England may move north to have their second brood, and juveniles marked in southern England have been found again in roosts in the north in the same season. Wood-pigeons also display northerly flights in September and October, before adopting a southerly orientation later in the autumn. Collared doves ringed in Europe as nestlings have moved north to Britain in the same autumn, and numbers of serins have turned up in south-west England in recent autumns. These movements seem adaptive in that young individuals which are surplus to the needs of the area in which they are born will stand more chance of finding new places to settle if they first explore north. The same principle applies to those birds of southeast and east Europe which might be expected to move north-west or west. I suspect that this factor may account for big arrivals of redbreasted flycatchers, woodchat shrikes, barred warblers, melodious and icterine warblers – all predominantly juvenile – into Britain in September 1958 and in subsequent years. Williamson showed that red-breasted flycatchers and icterine warblers arrived in Britain in clear anticyclonic weather with light winds, and as both migrate south-east to Asia, their movement several hundreds of miles off-course is remarkable. The explanation that they drifted in with down winds seems unlikely, and instead I wonder whether the existence of anticyclonic weather facilitated a normal adaptation after breeding in the form of a deliberate dispersal north-west, a process possibly truncated in years of less favourable weather.
Man has done so much in a passive way to alter the avifauna of Europe that it seems reasonable to take active steps to reintroduce lost species. Any reservations that this would be unnatural, should be tempered by the thought that the environment we have created is in any case artificial. Probably more pleasure than harm has been derived from the reintroduction of the capercaillie. It would seem laudable to follow up a recent suggestion and attempt the reintroduction of the bustard to parts of the East Anglian Breckland, and to encourage black terns to stay and breed. It is quite a different matter to introduce alien species to a new country, especially without sound biological knowledge. In Britain, some of these introductions, red-legged partridge, various pheasants, little owl, Canada and Egyptian goose and Mandarin duck have on balance improved our bird-life, but the same could not be said of the introduction of the house-sparrow and starling to Australia and North America.
Perhaps a more interesting question to ask here is why the majority of introduced species are unsuccessful. This is part of the much bigger question of what factors determine faunal diversity and enable some habitats to support more species than others. The concept of a niche, which refers to the animal’s place in the biotic environment and its relations to food and predators, should now be widely appreciated. It is a fundamental tenet of ecology that no two species can occupy the same niche in any one habitat, because both cannot be equally well adapted. R. and J. MacArthur (1961) examined numerous habitats at different latitudes for their plant species composition and foliage profile and ‘species diversity’. They found this last to be a more useful measure than the actual number of species because their calculations allowed at a habitat containing 50 of species A and 50 of B to rank a higher diversity than one with 99 of A and 1 of B. (The latter tends to be the farmland situation, the former that of tropical forest.) It turned out that neither the variety of plant species nor the latitude affected the amount of species diversity which instead depended entirely on the variation in foliage height, probably because birds mostly respond to different configurations of vegetation in different layers. This means that habitats of the same structural profile have the same diversity of bird species. In any area, a bird might either feed on all food of a suitable size within a narrowly defined habitat or, alternatively, be selective of food but collect it throughout a wider range of habitats. In other words, birds could partition their food or their habitat. The former has occurred because feeding specialisation brings the greatest advantages and has been favoured by natural selection. Partitioning the habitat would necessitate birds moving from one suitable micro-habitat (say a species of tree) to another, and it would depend on the pattern of the total habitat how much time would be wasted in the process. But adaptation to a comparatively broad habitat structure, for instance, to arboreal or ground feeding, must in turn impose physical limitations which restrict the diversity of feeding adaptations; in practice, bill size and shape is about all that can be much modified to suit the collection of different foods.
From the viewpoint of zoo-geography, the Palaearctic has existed as an entirety for sufficient time to ensure that most niches are filled by highly efficient species. Furthermore, man and birds have lived side by side since Neolithic times, so that the new habitats created by agriculture and man’s other activities have been occupied by the species best suited to them. The same is not true of Australia and New Zealand, which were cut off from the main centres of evolution at an earlier stage, one result being that primitive marsupial mammals were not replaced by the better adapted placental mammals. Birds are less insular, however, and the native avifauna of Australia seems to be the best fitted to occupy the niches available. Thus of at least 24 bird species deliberately introduced into Australia in the past, only 12 have become established. It is significant that only the blackbird has managed to invade native forest, the remainder existing in areas of recent agricultural development or urbanisation. But introduced birds like the feral pigeon, starling and house sparrow are better equipped to occupy the man-made niches than the native fauna, simply because these are species which have already been selected to occupy a man-made environment. New Zealand has an impoverished avifauna compared with Australia on which it has depended for colonisation, and the process is still incomplete. In consequence, fewer niches are saturated in New Zealand, so more exotic species have been successful. Of 130 species originally introduced, 24 have become established, although apart from the blackbird, chaffinch and redpoll which appear to be filling unexploited niches, most are again restricted to man-made habitats. Hawaii, which is even more isolated, and not saturated by a wide diversity of species, must have even more vacant niches, for, according to Elton (1958), of 94 birds introduced 53 have become established, some deep in the native forest.
According to Middleton, the European goldfinch has been successful in Australia and New Zealand only in the man-made agricultural areas, to which none of the native Australian Ploceid finches were adapted. In contrast, the European goldfinch has not been a successful bird in North America because it has virtually the same ecological requirements as the fitter endemic American goldfinch. Only one small colony of European goldfinches became established near New York, though these have since vanished when their habitat was destroyed for building purposes. Again the European house-sparrow has been highly successful in Australia, over roughly the same range as the goldfinch, whereas the introduced greenfinch is more restricted as it has rather more conservative ecotone requirements. It is interesting that this reflects a trend occurring today in Britain; the greenfinch is declining with the loss of hedgerows and woodland edges, while the goldfinch and linnet are increasing.
To return to New Zealand, it is noticeable that the birds which have become pests in agricultural areas, apart from being introduced species as one would expect from the comments above, present the same kinds of problems as they do in Britain. I am grateful to Dr P. C. Bull for allowing me to give details. As we shall see, skylarks (see here) are locally troublesome in Britain to young seedling crops such as lettuce. Near Hastings, N.Z., they and house sparrows have together been responsible for damaging asparagus and other seedlings. Both blackbird and song thrush and also the starling, resort to orchards in the dry season after breeding and cause considerable damage to all kinds of fruit, ripening pears, cherries and grapes. Redpolls do considerable damage to apricot blossom in their search for insects, and blossom searching is a habit which is increasing in Britain (see here). Locally in Britain, linnets peck out the seeds from strawberries (see here), while in N.Z. goldfinches do the same.
Various attempts were made to introduce the rook into N.Z. from 1762 onwards but only 35, liberated near Christchurch in 1873, seem to have thrived. The species occurs in five localities on the yellow-grey earths in the east of the county, generally where cereal growing occurs. Rooks at first increased very slowly but there was a rapid increase between 1935 and 1950, and a final levelling off with the density of birds in their favoured areas becoming virtually the same as that in Britain (around 16 nests per square mile). At Christ-church, the population increased from 1,000 birds in one rookery in 1925 to 7–10,000 in 1947 (13 rookeries), since when the numbers have remained roughly constant with 19 rookeries in use. Until 1926, the rookeries were in eucalypts, probably the favourite tree, but following a disease epidemic which killed these trees the birds changed to pines. Bull points out that the rate of increase of rooks has been slower than that of other introduced passerines and attributes this to their gregarious nesting habits and their need for group stimulation, and to an early shortage of suitable habitats. A feature of N.Z. rookeries is their very large size compared with British ones (rookeries of over 1,000 nests are quite common), and the traditional return to the same nesting sites may partly explain the slowness of expansion. (The large size of rookeries, and difficulties in getting sufficient food locally may also explain why the birds seem to lay, on average, smaller clutches in N.Z.; 3.4 eggs against 4 + in Britain, (see here), though more data are needed to establish the point.) Frequently, only when man actively disturbed these large rookeries did they become fragmented in surrounding areas, often with a rapid increase in total bird numbers in the district. As in Britain, rooks uproot seedling peas and corn, take ripening peas (and pumpkins) and maize and are also partial to walnuts.
The number of closely related birds which can live in the same habitat without competing for food depends to a large measure on the degree of stability within the environment. Marked fluctuations occur on English farmland, not only because of the changing seasons, but also because ploughing, harvesting and other farm operations impose drastic changes. As a result, the farmland birds occupying the various niches available for ground-feeders show a wide character displacement; we find a plover, three passerines (rook, starling, lark), a partridge and a pigeon, other species being only transient visitors, or primarily dependent on other habitats. No bird can afford to be too conservative in its niche requirements in a fluctuating environment, while the need for each species to show more tolerance reduces the number of ecologically isolated forms. Therefore, we should expect modern farm mechanisation, which enables whole farms to be ploughed within a fortnight, to be detrimental to bird life compared with the old methods which ensured some degree of stability by leaving land fallow and by transforming stubbles into bare ground more gradually. Klopfer and MacArthur (1960) have similarly emphasised that the major factor accounting for a decrease in the number of species away from the tropics, while the number of individuals of each species increases, does not result from a decrease in habitat complexity, but to a decrease in the similarity of coexisting species. The principle can obviously be extended to any situation where man simplifies the environment.
An important feature of complex ecological communities is that interactions between members damp out oscillations in the numbers of any one species (see here) and so help to introduce a high degree of stability and energy utilisation. For one thing, available food is more fully exploited, which is not the case in arctic environments for instance, where considerable seasonal changes occur. Hence, the amount of energy needed to maintain a stable community is less than that required for an unstable one. Man’s activities have tended to reduce complexity and introduce monotony, through monocultures of crops, uniform stands of trees, or rows of similar houses. In consequence, the animals inhabiting these environments usually fluctuate much more than those of more complex ecosystems, often to the extent of becoming pests (see here). One feature of stabilisation is that natural selection can favour anticipatory functions – for example, the breeding season of northern birds has become approximately geared to seasonal daylight changes – in unstable environments opportunism must set more of a premium. Because more energy goes into maintaining fluctuations in simple ecosystems, often short-term fluctuations, these systems offer more scope for rational exploitation, giving more production per unit of biomass. In general, pestiferous birds and game species can be cropped very intensively, but the corollary also applies in that the energy needed to counteract fluctuations, the efforts of pest control, must often be so considerable as to be impracticable. On the other hand, mature and complex ecosystems can be disturbed relatively easily. In the Eltonian food chain the predators at each level become rarer and larger, because the energy passing from link to link is only in the region of 10%-20%. This not only sets limits on the number of links in a food chain (five seems to be the maximum) and rules out the possibility of a super-predator but makes the top predators particularly vulnerable to small but cumulative changes in the food chain. The loss and increased rarity of so many of the birds of prey depends not so much on persecution, but on the reduced complexity of the environment through human ‘progress’. Clearly our future policies should not concentrate too much on bird protection per se, but rather on the creation and maintenance of as much diversified habitat as possible.
The long-term or ultimate value to a species in settling in an appropriate habitat will depend on the bird’s ability to find suitable food and produce surviving progeny, and this ability will be conditioned by the structural and behavioural adaptations of the species. The immediate or proximate factors which determine how a bird chooses an appropriate habitat are unlikely to involve these same factors. Instead, natural selection has enabled each species to respond to immediate signals, which can be reliably taken as indicators that other more basic needs will be satisfied. In this way a bird which lives in oak woodland might respond to the configuration of an oak tree, because natural selection will favour this appropriate response provided it leads the bird to find in the oak woods all the various foods which are appropriate to its needs and feeding adaptations. Natural selection can favour the emergence of appropriate proximate responses which are anticipatory.
In practice, it is generally agreed that birds respond to a range of releasing stimuli which combine to provide the best cues. A meadow pipit may respond innately to open country, thereafter to specific elements of the habitat, such as the height of grass, the presence of song posts and nest sites. These considerations are important when man radically alters the habitat, without necessarily altering its food value. As various features of the environment combine to produce a response, they need not always be present in the same proportions, and some may even be absent, for a response still to occur. Species vary in the capacity to respond when some stimuli are absent. Within any area intraspecific competition ensures that the most favourable sites are filled at the start, after which less complete habitats can be occupied. At low population densities, when, for example, a species is at the edge of its range, only the best habitats are occupied (stenotopy) whereas when population explosions occur, marginal areas are also utilised (eurytopy). This is well illustrated by the wide range of nesting sites accepted by the various gulls which have undergone a spectacular population explosion in Britain – nesting colonies occur on rocky and sandy sea shores, estuarine and freshwater marshes, inland lakes, and on moors and fells.
It becomes clear how an originally montane bird like the house martin should come to accept the sides of houses for its nest site instead of cliffs. Similarly the absence of a species from what appears to be a suitable habitat may be attributable to the absence of some apparently trivial factor which must be satisfied. Already in south Europe it is known that the presence of electric and telephone pylons and cables in otherwise open country facilitates colonisation by species like the collared dove and various shrikes.
The psychological response of the reed bunting to a limited range of habitat cues seems to have been the only reason for its past restriction to wetland habitats and its ecological isolation from the yellowhammer. But, as will emerge later, this segregation is no longer maintained and the invasion of yellowhammer habitats by the reed bunting is possibly the result of a genotypic change which has removed this psychological restriction. Another explanation is also tenable. Although most habitat recognition is innate, birds are able to reinforce or even modify, to a variable extent, these innate responses by learning processes. By this means, adults often return to traditional areas, even though these change drastically, whereas young birds breeding for the first time avoid entering habitats which do not release appropriate responses – either innate or acquired by imprinting in early life. For instance, Peitzmeier (1952) found that in a study area in Germany the curlew typically nested only in boggy areas and avoided surrounding cultivated land. When the marshes were drained and cultivated, the adults not only remained faithful to the area, but after learning its new characteristics, also spread to other tilled land which before they had avoided. It could be that this situation has applied in the case of the reed bunting. Peitzmeier attributes the further spread of curlews in arable environments to the imprinting of young which have been reared in these new habitats. Such processes explain why local populations of birds come to acquire unusual habitat associations – for example, the stone curlews which used to nest on the shingle of Dungeness and Norfolk beaches, or yellow wagtails which breed in fields of growing potatoes. Peitzmeier accounts for a remarkable and fairly sudden change in the nesting habitat of the mistle thrush in the same way – originally confined to continuous woodland dominated by conifers it began, in 1925, to nest in parkland-type habitat, and in small groups of deciduous trees in cultivated country. That very recent changes may have occurred in the habitat of an animal must always be remembered when trying to understand their adaptations – they may have evolved in conditions quite unlike those in which the birds are seen today.
CHAPTER 3
SOME PREDATORS AND THEIR PREY
THE last chapter was primarily concerned with the factors governing bird numbers and distribution, showing some of the ways in which man does or does not influence the natural balance. The importance of the food supply was stressed. In many cases the food supply can be considered as an independent variable. For example, the quantity of beech-mast varies in different years in response to climatic and other factors and is not initially determined by the birds which use it as food. Nevertheless, the availability of beech-mast profoundly affects the numbers of those species which have to depend on it for food. More bramblings winter in Britain in good beech-mast years. On the other hand, when short-eared owls feed on voles they may themselves become an important factor determining vole numbers. Watson (1893) quotes an interesting example chronicled for 1580. In that year a vole plague developed in the marshes near Southminster, Essex, which so depleted the grasses that cattle died and men were powerless to take any preventive action. The situation was supposedly saved by the arrival of ‘such a number of owls as all the shire was not able to yield; whereby the marsh-holders were shortly delivered from the vexations of the said mice’. When a reciprocal interaction exists between an animal and its food supply, it is termed a predator–prey relationship. The principles involved in such predator–prey interactions are fundamental to almost all aspects of economic ornithology, from the daily routine of the gamekeeper to the effects some forest birds may have on various insect pests. In discussing certain economically important examples it seems desirable to outline some of these principles.
