


Полная версия
Волосы. Иллюстрированное пособие для врачей, трихологов и парикмахеров


Фолликулярно-меланиновая единица
Активные меланоциты волосяной луковицы образуют функциональные единицы с соседними незрелыми кератиноцитами, которые получают гранулы меланина и образуют пигментированный стержень волоса. Меланин переносится преимущественно в кортекс стержня волоса, реже – в мозговой слой и лишь изредка – в кутикулу[39].
Фолликулярно-меланиновая единица состоит из меланоцитов и кератиноцитов волосяного матрикса, частично регулируемых фибробластами дермального сосочка. Пигментация – результат последовательных взаимодействий между этими клетками. Процесс пигментации включает в себя производство меланина меланоцитами, перенос гранул меланина в кортикальные и мозговые кератиноциты волоса и тем самым образование пигментированных стержней волос.
На каждые пять кератиноцитов в волосяной луковице в целом приходится один меланоцит. Меланоциты волосяной луковицы отличаются от эпидермальных тем, что они крупнее, имеют более разветвленные дендриты и более обширный аппарат Гольджи и эндоплазматический ретикулум. Они также производят более крупные меланосомы.
Синтез меланина и перенос пигмента в кератиноциты луковицы в значительной степени контролируются сигналами от кератиноцитов, иммунных клеток, фибробластов кожи и эндотелиальных клеток. Меланоциты могут влиять также и на окружающие клетки, например, с помощью переноса меланосом в кератиноциты или через производство и секрецию белков-регуляторов[40].
Примечание. Меланосома – это органелла, содержащая меланин и другие светопоглощающие пигменты.
Меланосомы
Меланины образуются в уникальных органеллах, называемых меланосомами. Эти связанные с лизосомами органеллы продуцируются в цитоплазме меланоцита в результате меланогенеза. Гранулы меланина транспортируются к периферии меланоцитов и в конечном итоге переносятся в соседние кератиноциты стержня волоса, за счет чего он пигментируется[41][42]. Ранние меланосомы продуцируются аппаратом Гольджи и возникают недалеко от ядра, а затем созревают до поздних (пигментированных) меланосом по мере продвижения к периферии меланоцита и его дендритам (отросткам).
Структура меланосом зависит от типа продуцируемого меланина. Меланоциты в темно-коричневых и черных волосяных фолликулах содержат наибольшее количество меланосом. Они называются истинными или эумеланосомами, а их меланин – истинным или эумеланином. Меланоциты в коричневой волосяной луковице также содержат эумеланосомы, но они несколько меньше по размеру, чем в черной. Меланоциты в луковицах светлых волосяных фолликулов производят только слабомеланизированные меланосомы с низким содержанием эумеланина. Меланоциты красных волосяных луковиц производят феомеланосомы с характерной неупорядоченной внутренней структурой. В отличие от фибриллярного ядра эумеланосомы, феомеланосома содержит везикулярный матрикс с красным и желтым меланином, расположенным нерегулярно в виде пятен и, по-видимому, случайным образом[43].
Меланогенез
Процесс меланогенеза можно разделить на две части: биогенез меланосом и работа биохимического пути, превращающего фенилаланин/1-тирозин в меланин. Оба процесса находятся под сложным генетическим контролем, при этом ферменты, структурные белки, факторы транскрипции, рецепторы и факторы роста, необходимые для этих процессов, кодируются более чем ста генами.
Давайте поговорим об основных белках, влияющих на выработку меланина в меланоцитах. К ним относятся:
• структурные белки меланосом, которые обеспечивают каркас для ферментов, необходимых для отложения меланина;
• белки, которые регулируют синтез меланина (три ключевых фермента) в меланосомах:
• TYR (тирозиназа),
• TYRP1 (родственный тирозиназе протеин-1),
• TRP2/DCT (допахромовая таутомераза).
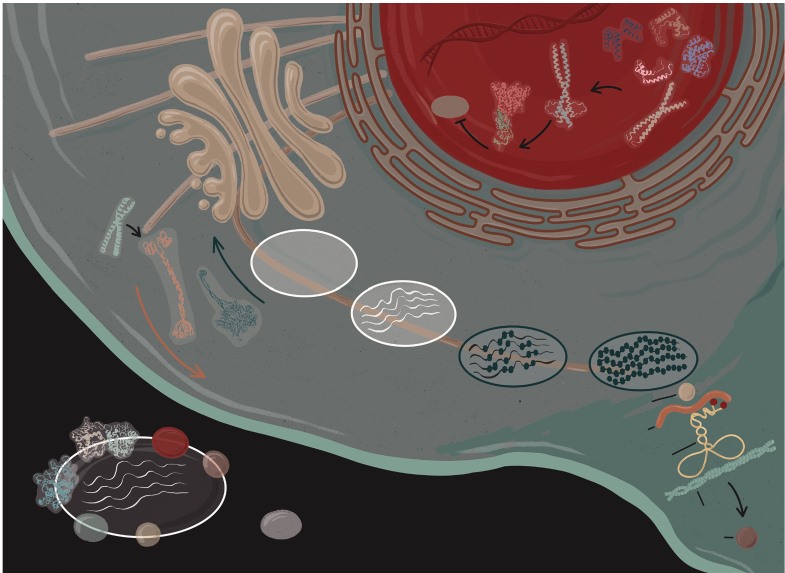
Рисунок 7. Факторы, регулирующие выработку меланина в меланоцитах. К ним относятся белки, которые влияют на структуру меланосомы (Pmel17, MART-1 и GPNMB); белки, которые прямо или косвенно модулируют синтез меланина (TYR, TYRP1, DCT, BLOC-1, OA1, P и SLC45A2), и белки, ответственные за транспортировку белков меланосом или внутриклеточный транспорт меланосом (микротрубочки, F-актин, кинезин, динеин, Rab27a, меланофилин, миозин Va, RILP, цилиобревины и Slp2-a), а также меланоцит-специфические факторы транскрипции (PAX3, SOX9/10, LEF-1, CREB, DICER и MITF). Стадии развития меланосом отмечены на этой диаграмме как I, II, III и IV.
На перемещение и функцию этих ферментов влияют многие факторы, включая BLOC-1, OA1, белок P и SLC45A2 (ранее известный как MATP). Стоит также отметить, что ионы металлов, включая цинк и медь, служат катализаторами работы фермента при синтезе меланина, а металлические остатки в окаменелостях позволяют даже предсказать цвет динозавров[44];
• белки, участвующие во внутриклеточном перемещении компонентов меланосом и транспорте меланосом к периферии клетки.
Цикличность меланогенеза волосяного фолликула
Наиболее важным различием между эпидермальными и фолликулярно-меланиновыми единицами является жесткий циклический контроль активности меланоцитов волосяной луковицы. Меланогенез в них активен только во время анагена. Эпидермальные меланоциты, напротив, кажутся все время функционирующими: наша кожа пигментирована постоянно. Для большинства людей эта активность может быть дополнительно стимулирована УФ-излучением[45]. Загар на коже – это не что иное, как эффект дополнительной стимуляции меланогенеза на солнце.
В волосяном фолликуле кожи головы человека период активного меланогенеза во время фазы анагена может длиться до 10 лет[46]. Он отключается во время перехода от анагена к катагену: сначала происходит подавление работы ключевых ферментов меланогенеза, за которым следует апоптоз меланоцитов волосяного фолликула[47].
Таким образом, фолликулярный меланогенез имеет характерную циклическую природу[48]. Ранние признаки неизбежной регрессии волосяного фолликула становятся очевидными в пигментной единице волосяного фолликула к концу анагена. К ним относятся втягивание дендритов меланоцитов и ослабление меланогенеза[49].
Во время телогена в волосяном фолликуле не вырабатывается меланин, но телогеновый волосяной фолликул все же содержит все клетки-предшественницы, нужные для восстановления полностью пигментированного волосяного фолликула во время анагена. Некоторые меланоцитарные клетки (вероятно, стволовые клетки меланоцитов) распределены по так называемым вторичным зародышевым областям телогеновых фолликулов. Они содержат кератиноциты и меланоциты, необходимые для начала следующего анагена.
Важный вопрос пигментации волосяного фолликула касается судьбы меланоцитов волосяной луковицы во время катагена: куда они уходят? И откуда они берутся, когда фолликулярный меланогенез возобновляется во время следующей фазы анагена? До недавнего времени господствовало мнение, что ниша меланоцитов волосяной луковицы самовоспроизводится. В соответствии с этой гипотезой меланоциты, участвующие в пигментации одного поколения волос, остаются вовлеченными в пигментацию следующего поколения, то есть существует несколько циклов де-дифференцировки меланоцитов с последующей повторной дифференцировкой.
Меланоциты в системе волосяных фолликулов более чувствительны к влиянию времени, чем меланоциты в эпидермисе, что приводит к поседению волос. Вероятно, это частично происходит из-за различий в эпидермальном и фолликулярном микроокружении клеток[50].
Возрастные изменения пигментации
Цвет волос, особенно у лиц евразийского происхождения, демонстрирует поразительные возрастные эффекты. В период полового созревания часто происходит переход от светлых «промежуточных волос» к более пигментированным и грубым «конечным волосам». Действительно, цвет волос у детей с возрастом обычно темнеет, и после полового созревания светловолосый ребенок нередко становится темноволосым.
По-видимому, с возрастом потеря активных меланоцитов происходит не только в коже (эпидермисе и волосяных фолликулах), но также в невусах и глазах. Уменьшение пигментации в эпидермисе происходит постепенно, а вот возрастная потеря цвета волос может быть гораздо более серьезной. Таким образом, разные субпопуляции меланоцитов регулируются разными «меланогенетическими часами».
Точный механизм потери меланоцитов как в эпидермисе, так и в волосяном фолликуле пока что неясен. Потеря способности меланоцитов активно делиться является, по-видимому, результатом их неспособности активировать путь митоген-активируемой протеинкиназы (MAPK), необходимый для активации их деления. При старении меланоциты также экспрессируют повышенные уровни ингибиторов циклин-зависимых киназ (например, p21 и p16), которые останавливают их клеточный цикл[51].
Вероятно, оказывает влияние и то, что антиоксидантные системы в меланоците волосяного фолликула с возрастом нарушаются, что приводит к неконтролируемому повреждению самого меланоцита, в том числе из-за его собственного окислительного стресса, связанного с меланогенезом. Уже известно, что поседение связано с усилением апоптоза меланоцитов и их окислительным стрессом[52][53]. Что касается экзогенного окислительного стресса, то из-за нарушения механизмов антиоксидантной защиты седеющие волосяные фолликулы менее приспособлены к борьбе с ним[54].
Стволовая ниша
Стволовые клетки
Взрослые стволовые клетки (СК) регулируют гомеостаз тканей и заживление ран. Они располагаются в специфической нише с особым микроокружением. СК могут самообновляться и оставаться недифференцированными на протяжении всей своей жизни[55][56]. В тканях они играют ключевую роль в регулировании гомеостаза и восстановлении повреждений[57]. Во время нормального гомеостаза они часто выходят из своих ниш и становятся транзитно-амплифицирующими (ТА) клетками, претерпевая серию быстрых делений перед окончательной дифференцировкой[58][59].
Взрослые СК имеют следующие характеристики:
• СК проживают в особой нише, важной для длительного поддержания их жизнедеятельности;
• СК редко делятся в течение жизни;
• при делении СК генерируют две дочерние клетки, одна из которых остается в нише и становится другой СК, а вторая покидает нишу и превращается в короткоживущую быстро делящуюся ТА клетку или клетку-предшественницу.
Важно подчеркнуть, что СК мультипотентны, то есть они способны генерировать все типы клеток своей собственной линии.
Выпуклость – стволовая ниша волосяного фолликула
Волосяные фолликулы содержат мультипотентные СК, которые активируются в начале нового цикла роста волос или при ранении для регенерации фолликула и восстановления эпидермиса.
Волосяной фолликул состоит из постоянной области (выпуклости), содержащей СК, и временной области (луковицы), содержащей дифференцированные клетки. Выпуклость – специализированная ниша, в которой находятся СК волосяного фолликула. Она расположена в его основании чуть ниже сальной железы – в том месте, где арректорная мышца прикрепляется к фолликулу. Выпуклость образуется во время морфогенеза эмбриона и приобретает свой характерный вид при появлении первого послеродового волосяного зародыша. Эта область не дегенерирует во время цикла роста волос.
В центре фолликула расположен стержень волоса, а СК локализуются в самом внешнем слое, который называется наружной корневой оболочкой. Выпуклость при этом является отдельной областью наружной корневой оболочки. В зрелых волосяных фолликулах наружная корневая оболочка простирается от выпуклости до матрикса.
Матрикс находится в основании луковицы. Это специализированный регион с клетками-предшественницами, которые быстро делятся и генерируют дифференцированные клетки, образующие внутреннюю корневую оболочку и стержень волоса. Он включает в себя скопление мезенхимальных клеток – дермальный сосочек[60].
В течение многих лет считалось, что СК, за счет которых во время цикла роста волос регенерирует фолликул, – это активно делящиеся матриксные клетки. Позднее, когда было обнаружено, что рентгеновское облучение убивает клетки матрикса, не нарушая при этом процесс формирования волос, такое мнение было подвергнуто сомнению.
Несмотря на то, что в целом клетки выпуклости находятся в состоянии покоя, их можно искусственно стимулировать к размножению в начале каждого цикла роста волос[61].
Рост волос происходит благодаря работе выпуклости следующим образом.
• СК активируются в начале анагена дермальным сосочком[62][63][64].
• После этого они мигрируют из выпуклости в матрицу вдоль наружной корневой оболочки.
• В матрице они размножаются и дифференцируются с образованием волос и внутренней корневой оболочки.
Выпуклость также является резервуаром мультипотентных СК, которые могут быть задействованы при заживлении ран, так как они участвуют в восстановлении эпидермиса[65].
В зоне выпуклости находятся, по крайней мере, две различные популяции СК различного происхождения,
• Мультипотентные СК кератиноцитов, отвечающие за циклическую регенерацию волосяных фолликулов, а также за поступление клеток в межфолликулярный эпидермис и сальные железы после ранения.
• СК меланоцитов, которые поставляют меланоциты, продуцирующие пигмент, в матрицу волос во время каждого цикла роста[66][67].
Следует отметить, что до сих пор нет ясности, в какой степени эти два разных типа СК взаимодействуют и способствуют функционированию друг друга[68].
СК обеспечивают непрерывный цикл жизни волосяного фолликула и его обновление, и они также являются источником эпителиальных клеток в процессе регенерации кожи. Зона выпуклости волосяного фолликула содержит несколько подгрупп стволовых клеток с различными функциями и регенеративным потенциалом. Другая область волосяного фолликула, перешеек, тоже содержит клетки со свойствами стволовых. Клетки перешейка участвуют в регенерации воронки (отверстия, из которого выходит волос) – она начинается на поверхности эпидермиса и продолжается до отверстия сального протока[69].
Адгезия в нише СК
Микроокружение и ниша играют решающую роль при поддержании пула СК. Адгезия клетки к клетке и клетки к внеклеточному матриксу в нише важна для создания и поддержания ее архитектуры[70].
Полудесмосомы представляют собой мультибелковые адгезионные комплексы, которые способствуют стабильному эпидермально-дермальному прикреплению. Трансмембранный белок коллаген XVII (COL17A1/BP180/BPAG2) – это структурный компонент полудесмосомной бляшки, которая выступает под полудесмосомами в базальных кератиноцитах эпидермиса в нижележащую базальную мембрану, таким образом скрепляя их друг с другом[71][72].
Белок COL17A1 участвует в гомеостазе волосяных фолликулов, высоко экспрессируется в СК внутри волосяных фолликулов и необходим для их самообновления. COL17A1-зависимое закрепление фолликулярных СК на базальной пластинке регулирует процессы чередования их состояния покоя и дифференцировки за счет изменения частоты их деления.
У пациентов с дефицитом COL17A1 полудесмосомы сформированы недостаточно эффективно, поэтому у таких пациентов преждевременно выпадают волосы и атрофируются волосяные фолликулы. Дефицит COL17A1 у мышей приводит к преждевременному и почти однородному выпадению волос по всей поверхности тела. Дефицит COL17A1 у человека также вызывает нелетальную форму буллезного соединительного эпидермолиза, также известную как генерализованный атрофический доброкачественный буллезный эпидермолиз. Выпадение волос у пациентов с буллезным эпидермолизом всегда связано с атрофией или потерей волосяного фолликула[73].
Изменения в выпуклости, связанные с циклом роста волос
Клетки зародыша волоса активируются в начале анагена. Перед этим СК находятся в длительном периоде отдыха, который длится месяцами[74][75]. Покоящиеся в выпуклости СК генерируют ранние клетки-предшественницы клеток волосяного фолликула. Последние, в свою очередь, производят быстро пролиферирующие клетки-предшественницы, расположенные в матриксе волоса. Впоследствии они окончательно дифференцируются и перемещаются вверх для формирования клонов внутренних волосяных фолликулов с внутренней корневой оболочкой и стержнем волоса. Матричные клетки подвергаются дифференцировке, а СК выпуклости проходят через короткую фазу самообновления в анагене, а затем снова возвращаются в состояние покоя[76]
Конец ознакомительного фрагмента.
Текст предоставлен ООО «Литрес».
Прочитайте эту книгу целиком, купив полную легальную версию на Литрес.
Безопасно оплатить книгу можно банковской картой Visa, MasterCard, Maestro, со счета мобильного телефона, с платежного терминала, в салоне МТС или Связной, через PayPal, WebMoney, Яндекс.Деньги, QIWI Кошелек, бонусными картами или другим удобным Вам способом.
Примечания
1
https://www.ncbi.nlm.nih.gov/books/NBK470321/
2
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
3
https://www.ncbi.nlm.nih.gov/books/NBK546248/
4
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7432488/
5
https://www.ncbi.nlm.nih.gov/books/NBK470321/
6
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7432488/
7
https://www.ncbi.nlm.nih.gov/books/NBK546248/
8
https://www.cell.com/current-biology/fulltext/S0960-9822(08)01626-6?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0960982208016266%3Fshowall%3Dtrue
9
https://www.ncbi.nlm.nih.gov/books/NBK546248/
10
https://www.ncbi.nlm.nih.gov/books/NBK470321/
11
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7432488/
12
https://www.cell.com/current-biology/fulltext/S0960-9822(08)01626-6?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0960982208016266%3Fshowall%3Dtrue
13
https://www.jidonline.org/article/S0022-202X(15)41553-2/fulltext
14
https://www.ncbi.nlm.nih.gov/books/NBK470321/
15
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2405915/
16
https://www.ncbi.nlm.nih.gov/labs/pmc/articles/PMC1175950/
17
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2871241/
18
https://pubmed.ncbi.nlm.nih.gov/10998156/
19
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1891949/
20
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2871241/
21
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2871241/
22
https://pubmed.ncbi.nlm.nih.gov/15617565/
23
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2756832/
24
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2871241/
25
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2756832/
26
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2668200/
27
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2871241/
28
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2705795/
29
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2871241/
30
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2871241/
31
https://www.jidonline.org/article/S0022-202X(15)41553-2/fulltext
32
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
33
https://www.cell.com/current-biology/fulltext/S0960-9822(08)01626-6?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0960982208016266%3Fshowall%3Dtrue
34
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3996377/
35
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
36
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
37
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
38
https://www.cell.com/current-biology/fulltext/S0960-9822(08)01626-6?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0960982208016266%3Fshowall%3Dtrue
39
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
40
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
41
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
42
https://www.cell.com/current-biology/fulltext/S0960-9822(08)01626-6?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0960982208016266%3Fshowall%3Dtrue
43
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
44
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3996377/
45
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
46
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
47
https://www.cell.com/current-biology/fulltext/S0960-9822(08)01626-6?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0960982208016266%3Fshowall%3Dtrue
48
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
49
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1201498/
50
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
51
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
52
https://pubmed.ncbi.nlm.nih.gov/16723385/
53
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
54
https://onlinelibrary.wiley.com/doi/10.1111/j.1468–2494.2008.00456.x
55
https://pubmed.ncbi.nlm.nih.gov/16810241/
56
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3050564/
57
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2756832/
58
https://pubmed.ncbi.nlm.nih.gov/16810241/
59
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3050564/
60
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2756832/
61
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2405915/
62
https://pubmed.ncbi.nlm.nih.gov/16288281/
63
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2756832/
64
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3050564/
65
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2405915/
66
https://pubmed.ncbi.nlm.nih.gov/11976685/
67
https://www.cell.com/cell-stem-cell/fulltext/S1934-5909(10)00651-X?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS193459091000651X%3Fshowall%3Dtrue
68
https://www.cell.com/cell-stem-cell/fulltext/S1934-5909(10)00651-X?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS193459091000651X%3Fshowall%3Dtrue
69
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7680886/