

 полная версия
полная версияAnts and Some Other Insects: An Inquiry Into the Psychic Powers of These Animals
The so-called reflexes and their temporary, purposefully adaptive, but hereditarily stereotyped combinations, which respond always more or less in the same manner to the same stimuli, constitute the paradigm of automatic activities. These have the deceptive appearance of a “machine” owing to the regularity of their operations. But a machine which maintains, constructs, and reproduces itself is not a machine. In order to build such a machine we should have to possess the key of life, i. e., the understanding of the supposed, but by no means demonstrated, mechanics of living protoplasm. Everything points to the conclusion that the instinctive automatisms have been gradually acquired and hereditarily fixed by natural selection and other factors of inheritance. But there are also secondary automatisms or habits which arise through the frequent repetition of plastic activities and are therefore especially characteristic of man’s enormous brain-development.
In all the psychic provinces of intellect, feeling, and will, habits follow the constant law of perfection through repetition. Through practice every repeated plastic brain-activity gradually becomes automatic, becomes “second nature,” i. e., similar to instinct. Nevertheless instinct is not inherited habit, but phylogenetically inherited intelligence which has gradually become adapted and crystalised by natural selection or by some other means.
Plastic activity manifests itself, in general, in the ability of the nervous system to conform or adapt itself to new and unexpected conditions and also through its faculty of bringing about internally new combinations of neurocyme. Bethe calls this the power of modification. But since, notwithstanding his pretended issue with anthropomorphism, he himself continually proceeds in an anthropomorphic spirit and demands human ratiocination of animals, if they are to be credited with plasticity (power of modification), – he naturally overlooks the fact that the beginnings of plasticity are primordial, that they are in fact already present in the Amœba, which adapts itself to its environment. Nor is this fact to be conjured out of the world by Loeb’s word “tropisms.”
Automatic and plastic activities, whether simple or complex, are merely relative antitheses. They grade over into each other, e. g., in the formation of habits but also in instincts. In their extreme forms they resemble two terminal branches of a tree, but they may lead to similar results through so-called convergence of the conditions of life (slavery and cattle-keeping among ants and men). The automatic may be more easily derived from the plastic activities than vice versa. One thing is established, however: since a tolerably complicated plastic activity admits of many possibilities of adaptation in the individual brain, it requires much more nervous substance, many more neurons, but has more resistances to overcome in order to attain a complicated result. The activities of an Amœba belong therefore rather to the plasticity of living molecules, but not as yet to that of coöperating nerve-elements; as cell-plasticity it should really be designated as “undifferentiated.”1 There are formed in certain animals specially complex automatisms, or instincts, which require relatively little plasticity and few neurons. In others, on the contrary, there remains relatively considerable nerve-substance for individual plasticity, while the instincts are less complicated. Other animals, again, have little besides the lower reflex centers and are extremely poor in both kinds of complex activities. Still others, finally, are rich in both. Strong so-called “hereditary predispositions” or unfinished instincts constitute the phylogenetic transitions between both kinds of activity and are of extraordinarily high development in man.
Spoken and especially written language, moreover, enable man to exploit his brain to a wonderful extent. This leads us to underestimate animals. Both in animals and man the true value of the brain is falsified by training, i. e., artificially heightened. We overestimate the powers of the educated negro and the trained dog and underestimate the powers of the illiterate individual and the wild animal.
I beg your indulgence for this lengthy introduction to my subject, but it seemed necessary that we should come to some understanding concerning the validity of comparative psychology. My further task now consists in demonstrating to you what manner of psychical faculties may be detected in insects. Of course, I shall select in the first place the ants as the insects with which I am most familiar. Let us first examine the brain of these animals.
In order to determine the psychical value of a central nervous system it is necessary, first, to eliminate all the nerve-centers which subserve the lower functions, above the immediate innervation of the muscles and sense-organs as first centers. The volume of such neuron-complexes does not depend on the intricacy of mental work but on the number of muscle-fibres concerned in it, the sensory surfaces, and the reflex apparatus, hence above all things on the size of the animals. Complex instincts already require the intervention of much more plastic work and for this purpose such nerve-centers alone would be inadequate.
A beautiful example of the fact that complex mental combinations require a large nerve-center dominating the sensory and muscular centers is furnished by the brain of the ant. The ant-colony commonly consists of three kinds of individuals: the queen, or female (largest), the workers which are smaller, and the males which are usually larger than the workers. The workers excel in complex instincts and in clearly demonstrable mental powers (memory, plasticity, etc.). These are much less developed in the queens. The males are incredibly stupid, unable to distinguish friends from enemies and incapable of finding their way back to their nest. Nevertheless the latter have very highly developed eyes and antennae, i. e., the two sense-organs which alone are connected with the brain, or supra-oesophageal ganglion and enable them to possess themselves of the females during the nuptial flight. No muscles are innervated by the supra-oesophageal ganglion. These conditions greatly facilitate the comparison of the perceptive organs, i. e., of the brain (corpora pedunculata) in the three sexes. This is very large in the worker, much smaller in the female, and almost vestigial in the male, whereas the optic and olfactory lobes are very large in the latter. The cortical portion of the large worker brain is, moreover, extremely rich in cellular elements. In this connection I would request you to glance at the figures and their explanation.
Very recently, to be sure, it has come to be the fashion to underestimate the importance of brain-morphology in psychology and even in nerve-physiology. But fashions, especially such absurd ones as this, should have no influence on true investigation. Of course, we should not expect anatomy to say what it was never intended to say.
In ants, injury to the cerebrum leads to the same results as injury to the brain of the pigeon.
In this place I would refer you for a fuller account of the details of sensation and the psychic peculiarities of insects to my more extended work above mentioned: Sensations des Insectes.
It can be demonstrated that insects possess the senses of sight, smell, taste, and touch. The auditory sense is doubtful. Perhaps a sense of touch modified for the perception of delicate vibrations may bear a deceptive resemblance to hearing. A sixth sense has nowhere been shown to occur. A photodermatic sense, modified for light-sensation, must be regarded as a form of the tactile sense. It occurs in many insects. This sense is in no respect of an optic nature. In aquatic insects the olfactory and gustatory senses perhaps grade over into each other somewhat (Nagel), since both perceive chemical substances dissolved in the water.
The visual sense of the facetted eyes is especially adapted for seeing movements, i. e., for perceiving relative changes of position in the retinal image. In flight it is able to localise large spatial areas admirably, but must show less definite contours of the objects than our eyes. The compound eye yields only a single upright image (Exner), the clearness of which increases with the number of facets and the convexity of the eye. Exner succeeded in photographing this image in the fire-fly (Lampyris). As the eyes are immovable the sight of resting objects soon disappears so far as the resting insect is concerned. For this reason resting insects are easily captured when very slowly approached. In flight insects orient themselves in space by means of their compound eyes. Odor, when perceived, merely draws these animals in a particular direction. When the compound eyes are covered, all powers of orientation in the air are lost. Many insects can adapt their eyes for the day or night by a shifting of the pigment. Ants see the ultra-violet with their eyes. Honey-bees and humble-bees can distinguish colors, but obviously in other tones than we do, since they cannot be deceived by artificial flowers of the most skilful workmanship. This may be due, to admixtures of the ultra-violet rays which are invisible to our eyes.
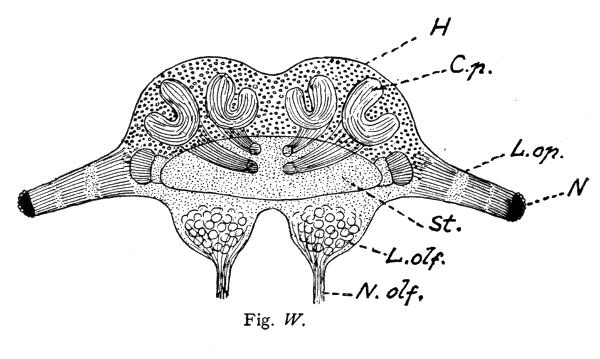
Fig. W.
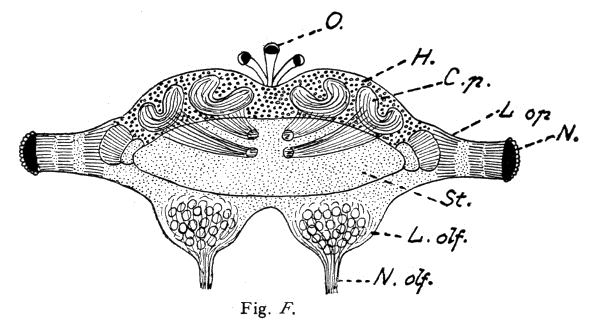
Fig. F.]
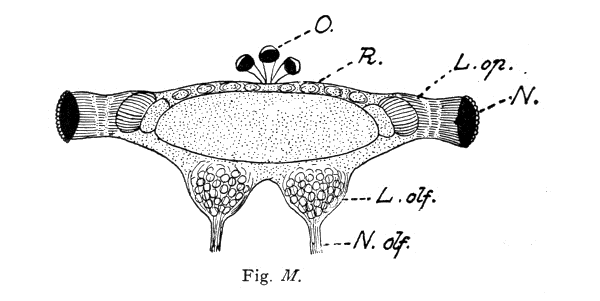
Fig. M.
EXPLANATION OF THE FIGURES
Brain (supra-œsophageal ganglion) of an ant (Lasius fuliginosus), magnified 60 diameters, seen from above.
Fig. W. Brain of the Worker.
Fig. F. Brain of the Queen (Female)
Fig. M. Brain of the Male.
St. = Brain trunk. L. op. = Lobus opticus (optic lobe). L. olf. = Lobus olfactorius sive antennalis (olfactory lobe). N. = Facetted eye. N. olf. = Nervus olfactorius sive antennalis (olfactory nerve). O. = Ocelli, or simple eyes with their nerves (present only in the male and queen). H. = Cellular brain cortex (developed only in the worker and queen). C. p. = Corpora pedunculata, or fungiform bodies (developed only in the worker and queen). R. = Rudimental cortex of male.
The length of the whole ant is:
in the worker 4.5 mm;
in the queen 6.0 mm;
in the male 4.5 mm.
N. B. The striation of the corpora pedunculata and their stems is represented diagrammatically, for the purpose of indicating rather coarsely their extremely delicate fibrillar structure.
The ocelli (simple eyes) play a subordinate rôle, and probably serve as organs of sight for objects situated in the immediate vicinity and in dark cavities.
The olfactory sense has its seat in the antennæ, usually in the club-shaped flagellum, or rather in the pore-plates and olfactory rods of these portions of the antennæ. On account of its external and moveable position at the tip of the antenna, the olfactory organ possesses two properties which are lacking in the vertebrates, and particularly in man. These are:
1. The power of perceiving the chemical nature of a body by direct contact (contact-odor);
2. The power of space-perception and of perceiving the form of objects and that of the animal’s own trail by means of odor, and the additional property of leaving associated memories.
The olfactory sense of insects, therefore, gives these animals definite and clear-cut perceptions of space-relations, and enables the animal while moving on the surface of the ground to orient itself with facility. I have designated this sense, which is thus qualitatively, i. e., in its specific energy, very different from our olfactory sense, as the topochemical (olfactory) sense. Probably the pore-plates are used for perceiving odor at a distance and the olfactory rods for contact-odor, but this is pure conjecture. Extirpation of the antennæ destroys the power of distinguishing friends from enemies and deprives the ant of the faculty of orienting itself on the ground and of finding its way, whereas it is possible to cut off three legs and an antenna without seriously impairing these powers. The topochemical sense always permits the ant to distinguish between the directions of its trail, a faculty which Bethe attributes to a mysterious polarisation. The ability to sense different odors varies enormously in different insects. An object possessing odor for one species is often odorless for other species (and for ourselves) and vice versa.
The gustatory organs are situated on the mouth-parts. Among insects the reactions of this sense are very similar to our own. Will accustomed some wasps to look in a particular place for honey, which he afterwards mixed with quinine. The wasps detected the substance at once, made gestures of disgust, and never returned to the honey. Mixing the honey with alum had the same result. At first they returned, but after the disagreeable gustatory experience they failed to reappear. Incidentally this is also a proof of their gustatory memory and of their powers of association.
Several organs have been found and described as auditory. But after their removal the supposed reaction to sounds persists. This would seem to indicate that a deceptive resemblance to hearing may be produced by the perception of delicate vibrations through the tactile sense (Dugès).
The tactile sense is everywhere represented by tactile hairs and papillæ. It reacts more especially to delicate tremors of the atmosphere or soil. Certain arthropods, especially the spiders, orient themselves mainly by means of this sense.
It may be demonstrated that insects, according to the species and conditions of life, use their different senses in combination for purposes of orienting themselves and for perceiving the external world. Many species lack eyes and hence also the sense of sight. In others, again, the olfactory sense is obtuse; certain other forms lack the contact-odor sense (e. g., most Diptera).
It has been shown that the superb powers of orientation exhibited by certain aerial animals, like birds (carrier-pigeons), bees, etc., depend on vision and its memories. Movement in the air gives this sense enormous and manifold values. The semi-circular canals of the auditory organ are an apparatus of equilibrium in vertebrates and mediate sensations of acceleration and rotation (Mach-Breuer), but do not give external orientation. For the demonstration of these matters I must refer you to my work above-cited. A specific, magnetic, or other mode of orientation, independent of the known senses, does not exist.
The facts above presented constitute the basis of insect psychology. The social insects are especially favorable objects for study on account of their manifold reciprocal relationships. If in speaking of their behavior I use terms borrowed from human life, I request you, once for all, to bear in mind that these are not to be interpreted in an anthropomorphic but in an analogous sense.
THE PROVINCE OF COGNITION
Many insects (perhaps all, in a more rudimental condition) possess memory, i. e., they are able to store up sense-impressions in their brains for subsequent use. Insects are not merely attracted directly by sensory stimuli, as Bethe imagines. Huber, myself, Fabre, Lubbock, Wasmann, Von Buttel-Reepen, have demonstrated this fact experimentally. That bees, wasps, etc., can find their way in flight through the air, notwithstanding wind and rain (and hence under circumstances precluding the existence of any possible odoriferous trail), and even after the antennæ have been cut off, to a concealed place where they have found what they desired, though this place may be quite invisible from their nest, and this even after the expiration of days and weeks, is a fact of special importance as proof of the above assertion. It can be shown that these insects recognise objects by means of their colors, their forms, and especially by their position in space. Position they perceive through the mutual relations and succession of the large objects in space, as these are revealed to them in their rapid change of place during flight in their compound eyes (shifting of retinal images). Especially the experiments performed by Von Buttel-Reepen and myself leave no doubt concerning this fact. Additional proof of a different nature is furnished by Von Buttel, who found that ether or chloroform narcosis deprives bees of all memory. By this means enemies can be converted into friends. Under these circumstances, too, all memory of locality is lost and must be reacquired by means of a new flight of orientation. An animal, however, certainly cannot forget without having remembered.
The topochemical antennal sense also furnishes splendid proofs of memory in ants, bees, etc. An ant may perform an arduous journey of thirty meters from her ruined nest, there find a place suitable for building another nest, return, orienting herself by means of her antenna, seize a companion who forthwith rolls herself about her abductrix, and is carried to the newly selected spot. The latter then also finds her way to the original nest, and both each carry back another companion, etc. The memory of the suitable nature of the locality for establishing a new nest must exist in the brain of the first ant or she would not return, laden with a companion, to this very spot. The slave-making ants (Polyergus) undertake predatory expeditions, led by a few workers, who for days and weeks previously have been searching the neighborhood for nests of Formica fusca. The ants often lose their way, remain standing and hunt about for a long time till one or the other finds the topochemical trail and indicates to the others the direction to be followed by rapidly pushing ahead. Then the pupæ of the Formica fusca nest, which they have found, are brought up from the depths of the galleries, appropriated and dragged home, often a distance of forty meters or more. If the plundered nest still contains pupæ, the robbers return on the same or following days and carry off the remainder, but if there are no pupæ left they do not return. How do the Polyergus know whether there are pupæ remaining? It can be demonstrated that smell could not attract them from such a distance, and this is even less possible for sight or any other sense. Memory alone, i. e., the recollection that many pupæ still remain behind in the plundered nest can induce them to return. I have carefully followed a great number of these predatory expeditions.
While Formica species follow their topochemical trail with great difficulty over new roads, they nevertheless know the immediate surroundings of their nest so well that even shovelling away the earth can scarcely disconcert them, and they find their way at once, as Wasmann emphatically states and as I myself have often observed. That this cannot be due to smelling at long range can be demonstrated in another manner, for the olfactory powers of the genus Formica, like those of honey-bees, are not sufficiently acute for this purpose, as has been shown in innumerable experiments by all connoisseurs of these animals. Certain ants can recognise friends even after the expiration of months. In ants and bees there are very complex combinations and mixtures of odors, which Von Buttel has very aptly distinguished as nest-odor, colony- (family-) odor, and individual odor. In ants we have in addition a species-odor, while the queen-odor does not play the same rôle as among bees.
It follows from these and many other considerations that the social Hymenoptera can store up in their brains visual images and topochemical odor-images and combine these to form perceptions or something of a similar nature, and that they can associate such perceptions, even those of different senses, especially sight, odor, and taste, with one another and thereby acquire spatial images.
Huber as well as Von Buttel, Wasmann, and myself have always found that these animals, through frequent repetition of an activity, journey, etc., gain in the certainty and rapidity of the execution of their instincts. Hence they form, very rapidly to be sure, habits. Von Buttel gives splendid examples of these in the robber-bees, i. e., in some of the common honey-bees that have acquired the habit of stealing the honey from the hives of strangers. At first the robbers display some hesitation, though later they become more and more impudent. But he who uses the term habit, must imply secondary automatism and a pre-existing plastic adaptability. Von Buttel adduces an admirable proof of this whole matter and at the same time one of the clearest and simplest refutations of Bethe’s innumerable blunders, when he shows that bees that have never flown from the hive, even though they may be older than others that have already flown, are unable to find their way back even from a distance of a few meters, when they are unable to see the hive, whereas old bees know the whole environment, often to a distance of six or seven kilometers.
It results, therefore, from the unanimous observations of all the connoisseurs that sensation, perception, and association, inference, memory and habit follow in the social insects on the whole the same fundamental laws as in the vertebrates and ourselves. Furthermore, attention is surprisingly developed in insects, often taking on an obsessional character and being difficult to divert.
On the other hand, inherited automatism exhibits a colossal preponderance. The above-mentioned faculties are manifested only in an extremely feeble form beyond the confines of the instinct-automatism stereotyped in the species.
An insect is extraordinarily stupid and inadaptable to all things not related to its instincts. Nevertheless I succeeded in teaching a water-beetle (Dytiscus marginalis) which in nature feeds only in the water, to eat on my table. While thus feeding, it always executed a clumsy flexor-movement with its fore-legs which brought it over on its back. The insect learned to keep on feeding while on its back, but it would not dispense with this movement, which is adapted to feeding in the water. On the other hand, it always attempted to leap out of the water (no longer fleeing to the bottom of the vessel) when I entered the room, and nibbled at the tip of my finger in the most familiar manner. Now these are certainly plastic variations of instinct. In a similar manner some large Algerian ants which I transplanted to Zurich, learned during the course of the summer months to close the entrance of their nest with pellets of earth, because they were being persecuted and annoyed by our little Lasius niger. In Algiers I always saw the nest-opening wide open. There are many similar examples which go to show that these tiny animals can utilise some few of their experiences even when this requires a departure from the usual instincts.
That ants, bees, and wasps are able to exchange communications that are understood, and that they do not merely titillate one another with their antennæ as Bethe maintains, has been demonstrated in so many hundred instances, that it is unnecessary to waste many words on this subject. The observations of a single predatory expedition of Polyergus, with a standing still of the whole army and a seeking for the lost trail, is proof sufficient of the above statement. But, of course, this is not language in the human sense! There are no abstract concepts corresponding to the signs. We are here concerned only with hereditary, instinctively automatic signs. The same is true of their comprehension (pushing with the head, rushing at one another with wide-open mandibles, titillation with the antennæ, stridulatory movement of the abdomen, etc.). Moreover, imitation plays a great rôle. Ants, bees, etc., imitate and follow their companions. Hence it is decidedly erroneous (and in this matter Wasmann, Von Buttel, and myself are of but one opinion) to inject human thought-conception and human ratiocination into this instinct-language, as has been done to some extent, at least, even by Pierre Huber, not to mention others. It is even very doubtful whether a so-called general sensory idea (i. e., a general idea of an object, like the idea “ant,” “enemy,” “nest,” “pupa”) can arise in the emmet brain. This is hardly capable of demonstration. Undoubtedly perception and association can be carried on in a very simple way, after the manner of insects, without ever rising to such complex results. At any rate proofs of such an assumption are lacking. But what exists is surely in itself sufficiently interesting and important. It gives us at least an insight into the brain-life of these animals.