




Полная версия
Эволюция. От Дарвина до современных теорий



Как именно возникло это новое понимание? И почему потребовалось так много времени?
Объяснение начинается с самого естественного отбора. В соответствии с концепцией естественного отбора, выживают и размножаются только самые приспособленные к локальной среде обитания. Таким образом, популяция как единое целое постепенно изменяется. Сама идея эволюции была принята многими биологами еще в середине XIX века. Однако при попытках признать тот факт, что эволюция происходит под влиянием естественного отбора, возникли некоторые разногласия.
Вероятность существования данного механизма базируется на предположении о том, что благоприятные признаки передаются от одного поколения к следующему в более-менее неизменном виде. Однако никто не знал, как именно это происходит.
Объяснение наследованияДарвин пытался объяснить наследственность с точки зрения гипотезы под названием «пангенезис». Ученый считал, что каждый организм производит определенные частицы, называемые геммулами, которые передают свои признаки следующему поколению. Дарвин предположил, что потомство развивается из сочетания родительских геммул, благодаря чему демонстрируется комбинация их черт.
Но у этой идеи был серьезный недостаток, которым пользовались его оппоненты: сочетание признаков привело бы к «разбавлению» полезных признаков одного из родителей при скрещивании с особями, не обладающими данными признакам. На протяжении следующих поколений такие признаки должны были неизбежно исчезнуть. И при жизни Дарвина никто не мог решить данную проблему.
Однако ключ к разгадке был уже известен, о чем не знал ни Дарвин, ни его соотечественники. В 1840-х годах Грегор Мендель пришел в мужской монастырь в Брно (Чешская Республика). За последующие годы он провел подробные исследования того, как из поколения в поколение передавались определенные признаки гороха. Монах заметил, что признаки родителей у потомков не смешивались. Скорее, они передавались без изменений, подчиняясь неким закономерностям. Это подтолкнуло Менделя к разработке законов наследования, опубликованных в 1866 году (см. «Кем был Грегор Мендель?»).
Однако никто и не предполагал, что признаки, изучаемые Менделем в горохе (окрас цветка), будут иметь столь важное значение. И на протяжении десятилетий работа исследователя игнорировалась.
Кем был Грегор Мендель?
Жизненный путь Грегора Менделя был крайне необычным для человека, считающегося основателем современной генетики, хотя бы потому, что он провел свою исследовательскую работу за 50 лет до фактического обнаружения генов. Мендель родился в 1822 году на ферме, находящейся на территории современной Чешской Республики. Затем он ушел в мужской монастырь в Брно, где начал заниматься изучением наследственности.
В монастырском саду он вырастил тысячи растений гороха, отметив наличие у них таких признаков, как окрас цветка и морщинистость семян. Например, он обнаружил, что при скрещивании белоцветных растений с фиолетовыми окрас получался не светло-лиловым (как это предписывала концепция о смешении родительских черт), а либо белым, либо фиолетовым в определенном соотношении.

Рис. 3.1. Жизненный путь Грегора Менделя был крайне необычным для человека, ставшего основателем современной генетики.
Эти наблюдения привели его к разработке известных нам законов наследования, опубликованных в 1866 году и представлявших идею доминантных и рецессивных признаков. Данная работа оставалась незамеченной до следующего столетия, когда идеи Менделя были включены в новое научное направление – генетику.
С однозначными признанием вклада Менделя в развитие этой науки согласны не все. Его законы, безусловно, пролили свет на механизм передачи признаков от родителей к потомству. Но даже Рональд Фишер, использовавший идеи Менделя для создания генетической теории эволюции в 1930-х годах, признавал, что результаты Менделя оказались слишком хороши для того, чтобы быть правдой, и, возможно, были «приукрашены» чрезмерно ретивым ассистентом. При этом ни в коем разе нельзя утверждать, что сам Мендель стал бы сторонником теории, основанной на его работе. Свои идеи он выказывал исключительно с точки зрения передачи признаков от одного поколения к другому, не обсуждая при этом сам механизм.
Со временем Мендель забросил свои исследования, став аббатом в возрасте 46 лет. О нем мало что известно, так как его переписка и прочие личные документы были сожжены после его смерти.
Затем, в 1900 году, законы Менделя заново открыли ботаники Хуго Де Фриз и Карл Корренс. Изучая наследование, оба ученых, независимо друг от друга, пришли к единому мнению о том, что признаки организма являются структурированными единицами и передаются следующему поколению без изменений. И только позже Де Фрис и Корренс обнаружили, что те же исследования проводились Менделем.
Расцвет геновТак зародилась новая наука о наследовании, которую вначале окрестили «менделизмом». Затем она была переименована в «генетику» биологом Уильямом Бэтсоном, который перевел статью Менделя на английский язык и стал главным пропагандистом его работы. Название для нового научного направления Бэтсон образовал из древнегреческого слова «генезис», что означает «происхождение».
Мендель выражал свои законы с точки зрения признаков, передаваемых родителем потомку. Ранние генетики полагали, что некоторая материальная сущность в организме должна была закодировать эту информацию.
Вскоре биолог Томас Хант Морган определил гены как структуры, расположенные вдоль хромосом внутри клеточного ядра. Работая над плодовой мушкой дрозофилой в 1910 году, Морган показал, что признак, отвечающий за цвет глаз, можно проследить до определенного отрезка Х-хромосомы. Это привело к революционным открытиям связей между различными генами и созданию генетических карт, показывающих расположение генов на хромосомах.
В конце концов исследования Моргана принесли ему Нобелевскую премию и подтвердили идею о том, что гены являются физической сутью наследования. Однако потребовалось еще целых три десятилетия, чтобы обнаружить, что гены состоят из ДНК и что каждый ген кодирует определенный белок.
Казалось, что концепция генов и была той самой недостающей частью теории Дарвина. Она дорисовывала картину естественного отбора, показывая, что признаки не могут смешиваться до неузнаваемости. Стоит добавить, что данный факт был признан не сразу.
Генетика решала еще одну проблему теории Дарвина – источник вариаций в популяции. Отправной точкой Дарвиновской теории служил тот факт, что по природе своей любая популяция содержит множество особей, обеспечивающих исходный материал для естественного отбора.
Теперь же было доказано, что основным источником этого разнообразия служат мутации – спонтанные изменения в структуре гена, приводящие к тому, что он начинает кодировать нечто новое. Такие изменения отмечались Морганом и другими исследователями при изучении положения генов в хромосомах.
Сам Морган пришел к выводу, что вредные мутации быстро искореняются из популяции, признавая тем самым отрицательную сторону естественного отбора. Однако для демонстрации положительного эффекта естественного отбора на гены требовалась дополнительная работа.
Быстро или медленно?По мнению Дарвина, эволюция – это медленный процесс постепенной адаптации к среде, в ходе которой большинство признаков имеют или когда-либо имели адаптивную функцию. Жирафы с чуть более длинными шеями могли дотянуться до более высоко расположенных листьев, поэтому в процессе естественного отбора у животных появлялись более длинные шеи. Большинство ранних генетиков, напротив, рассматривали эволюцию как нечто, происходящее большими скачками (или сальтациями), в результате чего новые признаки неожиданно проявлялись в ходе какой-либо внутренней перестройки наследственной конституции организма. Например, на растении могли внезапно появиться цветы другого окраса, отличного от родительского. Подобное изменение не обязательно будет носить адаптивный характер.
Ранних генетиков привлекали законы Менделя из-за кажущейся поддержки данных идей. Морган считал, что «природа создает новые виды сразу идеальными» посредством «внезапного изменения зародыша». Бэтсон не видел никакой ценности в исследованиях дарвинистов о непрерывной изменчивости и не согласился с утверждением, что естественные признаки появляются в результате адаптивного давления на виды. По той же самой причине сальтационный механизм изменений не имел никакого отношения к процессу естественного отбора.
Эти укоренившиеся взгляды не давали возможности найти способ согласования двух подходов. Но ситуация изменилась в 1920-х годах благодаря новой области популяционной генетики – изучению изменений определенных генов внутри популяций в течение времени.
Биологи Рональд Фишер, Джон Бердон Сандерсон Холдейн и Сьюалл Райт задействовали сложные математические модели, чтобы показать, что естественный отбор способен увеличить частоту кодирования любого гена для полезного признака и избавиться от наборов генов, не несущих адаптивной нагрузки.
Эта концепция была разработана в книге Фишера «Генетическая теория естественного отбора» (1930) и наиболее известной книге Холдейна «Причины эволюции» (1932). В том же году Райт представил идею адаптивного ландшафта – карты, изображающей все возможные комбинации генов и общей приспособляемости организма.
В итоге работа этих ученых доказала, что гены лежат в основе как резких изменений признаков, периодически встречаемых у потомства, так и постоянной изменчивости, зафиксированной Дарвином у больших популяций. Биологи показали, что генетический отбор является крайне изобретательной силой, руководящей адаптацией вида к локальной среде, с постоянной мутацией для поддержания генофонда изменчивости. Однако их теоретические модели основывались на сложной статистике и были сложны для понимания.
Научное сообщество познакомилось с принципами геноцентризма в эволюции лишь в 1937 году, когда Феодосий Добржанский опубликовал свою книгу «Генетика и происхождение видов», переведя в ней математические формулы на простой язык. Работа Добржанского расширила наше понимание того, как генетика способствует эволюции, показав, как в результате изменчивости для адаптации к локальной среде происходит образование новых видов у изолированных популяций.
В 1942 году биолог Джулиан Хаксли в своем детальном обзоре «Эволюция: современный синтез» дал название этому новому направлению. К 1950-м годам стала доминировать его формулировка эволюции, однако один из самых ключевых аспектов теории так и остался неоднозначным на протяжении следующих десятилетий.
От генетики к евгенике
Некоторые ранние приверженцы эволюционной теории являлись восторженными сторонниками евгеники – идеи увеличения человеческой популяции путем устранения «непригодных» генов.
К примеру, Рональд Фишер посвятил часть своей книги «Генетическая теория естественного отбора» (1930) надеждам на улучшение человеческой расы за счет евгеники. Чтобы еще активнее поспособствовать реализации данных принципов, он стал отцом восьмерых детей.
Обратная сторона евгеники дала о себе знать в начале XX века, когда несколько штатов США приняли закон о стерилизации «умственно отсталых», а нацисты довели эту идею до радикальной и ужасающей крайности.
Высшая цель?С момента своего зарождения дарвиновская теория эволюции так или иначе рассматривалась как идея, идущая в разрез с христианским видением природы как продукта некой высшей цели. Наиболее распространенной областью для креационистской оппозиции дарвиновской теории стала территория США, расцвет ее приходится на 1920-е годы и продолжается до сих пор.
Основатели «современного синтеза» хотели показать, что дарвинизм способен сочетать в себе веру в то, что для эволюции характерна тенденция к созданию более высоких уровней организации. Добржанский был выходцем из русской православной среды и в 1962 году написал книгу «Эволюционирующее человечество», пытаясь донести свою мысль о том, что эволюция имеет конечную цель. Хаксли также активно продвигал идею эволюционного прогресса.
Эти авторы представили идеи современного синтеза таким образом, что они не так открыто противоречили традиционным устоям и ценностям. Однако это не помешало некоторым ученым стать сторонниками евгеники (см. «От генетики к евгенике» выше). В последующие десятилетия псевдорелигиозный подход сойдет на нет и поспособствует этому публикация книги «Эгоистичный ген» (1976) Ричарда Докинза, в которой автор позиционирует себя в качестве главного приверженца идеи о том, что природа не имеет никакой конечной моральной цели. Последующие дебаты об эволюции социального поведения и появлении альтруизма проходили на фоне растущей напряженности между дарвиновской эволюцией и религией, то есть всего того, что основатели синтеза надеялись избежать.
Рис. 3.2. Двойная спираль ДНК была открыта в 1953 году.
Современный синтез, несмотря на все сопутствующие трудности, так и остался лежать в основе нашего понимания эволюции. Само понимание эволюции развивается по мере того, как новые открытия в генетике, онтогенетике и экологии расширяют наше понимание взаимосвязей между генами, организмами и окружающей средой.
Геноцентричный взгляд на эволюцию, ответвившийся от идей Дарвина и Менделя, также изменился после того, как мы признали, что среда развития организма играет определенную роль в формировании признаков и способна влиять на способ передачи этих признаков будущим поколениям.
Открытия в области эпигенетики показывают, что химические метки, прикрепляемые для включения/выключения генов, могут оказаться столь же важными для развития, как и сам запрограммированный генетический код (см. главу 8).
Современный синтез считается идеей ХХ века. В ХХI веке история эволюции обретет тот самый уровень совершенства, о котором Дарвин мог только мечтать (см. главу 11).
Что же такое «ген»?
В большинстве своем ген состоит из последовательности ДНК, кодирующей белок, и регуляторных последовательностей (промоторов), которые определяют, когда, где и сколько белка производить. В сложных клетках кодирующая последовательность делится на несколько частей (экзонов), которые отделяются более длинными участками «мусорной» ДНК (интронами) (см. рис. 3.3).
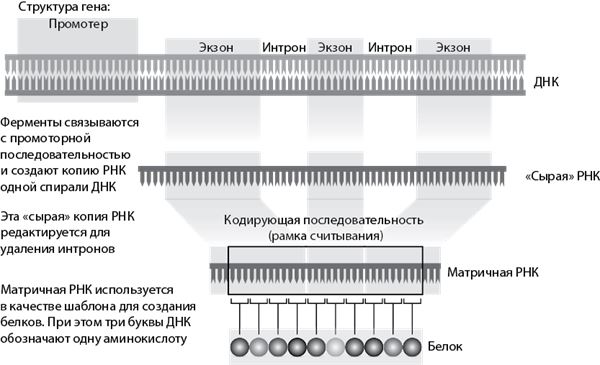
Рис. 3.3. Что такое ген?
Как происходит эволюция генов
Благодаря секвенированию геномов все большего количества видов, мы можем не только проследить за эволюцией тел животных, но и определить генетические мутации, стоящие за подобными изменениями.
Но самое интересное здесь то, что теперь мы можем увидеть, как возникают гены – главные составляющие ключевой основы жизни – белков. И история разворачивается не совсем так, как ожидалось.
Самый очевидный путь развития нового гена – это постепенное накопление незначительных полезных мутаций. Наименее очевидный сценарий: существующий ген, играющий важную роль, эволюционирует в другой ген. Вероятность того, что уже существующий ген сможет развиться в новый без изменения самого организма, весьма мала. Однако, как было замечено биологами столетие назад, данное ограничение вполне преодолимо в случаях, когда мутации создают дополнительную и полноценную копию гена.
Из учебников мы знаем, что процесс формирования новых генов начинается с дупликации генов. В большинстве случаев одна из копий генов приобретет вредные мутации и будет отсеяна. Однако иногда случается и так, что мутация позволяет дублированному гену выполнять нечто новое. Данная копия становится специализированной для своей новой роли, а предковый ген продолжает выполнять старые функции.
Поразительно то, что дупликация генов оказалась почти так же распространена, как и мутации, которые изменяют одну «букву» кода ДНК. При обмене материалом между хромосомами, предшествующему половому размножению, ошибки могут создавать дополнительные копии длинных последовательностей ДНК, в которых может содержаться любая информация – от одного гена до сотен. Здесь, как и при синдроме Дауна, могут дублироваться целые хромосомы, а иногда даже целые геномы.
Поскольку в процессе эволюции дублирование способно создать триллионы генетических копий, нет ничего удивительного в том, что в течение сотен миллионов лет один предковый ген способен породить сотни новых. У нас, людей, имеется порядка 400 генов для одних только обонятельных рецепторов. И все они происходят всего от двух рыб, живших около 450 миллионов лет назад.
Не конец историиИ все же классические взгляды на эволюцию генов не дают объяснения всей картины в целом. Зачастую гены несут в себе более одной функции… так что же происходит при дублировании гена? Если мутация убирает одну из двух существующих функций в первой копии гена, то организм сможет прекрасно существовать, поскольку вторая копия останется неизменной. Даже если еще одна мутация уберет другую функцию из второй копии гена, организм продолжит свое нормальное функционирование.
Теперь вместо одного гена с двумя функциями организм получит два гена с одной функцией в каждом. Данный механизм получил название «субфункционализация». Этот процесс может служить исходным материалом для дальнейшей эволюции.
Но настоящая проблема классической модели эволюции обуславливается фактическими исследованиями новых генов в различных организмах. Например, сравнение геномов нескольких близкородственных видов дрозофилы обнаружило новые гены, которые образовались через 13 миллионов лет после отделения данного вида от общего предка.
Так стало ясно, что около 10 % новых генов возникает в результате процесса под названием «ретропозиция». Ретропозиция происходит в тот момент, когда копии генов матричной РНК (генетические схемы, отправляемые на фабрику по производству белка в клетке, см. рис. 3.3) возвращаются обратно в ДНК, которая затем внедряется в другом месте генома.
Многие вирусы и генетические паразиты копируют себя с помощью ретропозиции, а производимые ими ферменты иногда случайным образом ретропозицируют РНК клетки-хозяина.
Возможно, что этот процесс ответственен за создание многих из недавно появившихся генов у нас, приматов. Вспышка ретропозиции у наших предков, достигшая своего пика около 45 миллионов лет назад, дала толчок к развитию многих тысяч дубликатных генов, и около 60–70 из них эволюционировали в новые гены. Данная вспышка, скорее всего, была обусловлена появлением нового генетического паразита, проникавшего в наш геном.
Эволюция новых генов часто включает в себя еще более радикальные изменения. Например, у дрозофил треть новых генов значительно отличалась от родительских, теряя часть своих последовательностей или приобретая новые участки ДНК.
Откуда берутся эти дополнительные последовательности? В сложных клетках ДНК, кодирующая белок, разбивается на несколько частей, разделенных некодирующими последовательностями. После создания РНК-копии всего гена некодирующие части (интроны) вырезаются, а кодирующие части (экзоны) сращиваются. Затем эта измененная копия РНК отправляется на белковую фабрику. Модульная форма генов значительно увеличивает вероятность мутаций благодаря перетасовке существующих генов и генерации новых белков. Происходить это может различными способами: экзоны внутри гена могут теряться, дублироваться или даже объединяться с экзонами других генов для создания нового химерного гена.
Вариации на темуНапример, большинство обезьян производят белок под названием TRIM5, который защищает их от заражения ретровирусами. Около 10 миллионов лет назад у одной макаки из Азии рядом с геном TRIM5 была добавлена неактивная копия гена CypA, полученная путем ретропозиции. Дальнейшая мутация привела к тому, что клетки продуцировали химерный белок, который на половину TRIM5, а на половину – CypA.
Данный белок обеспечивал лучшую защиту от некоторых вирусов. В это сложно поверить, но ген TRIM5-CypA эволюционировал не один, а целых два раза. Почти то же самое произошло с трехполосыми дурукули в Южной Америке.
При наличии достаточного количества времени – или, скорее, достаточного количества мутаций – дупликация и перетасовка генов может приводить к появлению новых генов, значительно отличающихся от предковых. Но все ли новые генные вариации соответствуют друг другу или же эволюция способна создавать новые гены, отличные от уже существующих?
Пару десятилетий назад было высказано предположение о том, что уникальные гены могут возникать в результате так называемой мутации сдвига рамки считывания. Каждая аминокислота в белке определяется тремя «буквами» ДНК или нуклеотидами – триплетом (кодоном). Если мутация сдвигает начальную точку считывания кодонов (рамку считывания) на один или два нуклеотида, то конечная последовательность белка будет совершенно иной.
Поскольку ДНК состоит из двух цепочек, то любой ее фрагмент можно «прочитать» шестью различными способами.
Генетический абсурдПодавляющее большинство мутаций, изменяющих рамку считывания гена, приводят к появлению генетического абсурда. Как правило, опасного. Многие генетические заболевания являются результатом мутации сдвига рамки считывания, разрушающей белки. Это немного похоже на замену каждой буквы алфавита на соседнюю. Результат, как правило, получается абсурдным. Но не всегда.
Другим источником уникальных новых генов может быть «мусорная» ДНК, засоряющая большинство геномов. Первые догадки об этом были высказаны два десятилетия назад, когда команда из Иллинойского университета раскрыла происхождение антифризного белка, вырабатываемого одной антарктической рыбой. Изначально данный ген появился в качестве пищеварительного фермента. Около 10 миллионов лет назад, когда климат на планете стал прохладнее, часть одного из интронов (иными словами – часть «мусорной» ДНК) превратилась в экзон, а затем многократно дублировалась, создавая характерную повторяющуюся структуру антифризных белков.
Так из случайного фрагмента ДНК развился ген, жизненно необходимый для выживания рыбы. Тем не менее этот антифризный ген эволюционировал из уже существующего.
Каковы шансы появления мутаций в «мусорной» ДНК, которые смогли бы сгенерировать полноценный новый ген с нуля? Как до недавнего времени считало большинство биологов, – практически нулевые. Ведь для того, чтобы фрагмент случайной ДНК превратился в ген, потребуется целый комплекс маловероятных условий. Во-первых, некая часть ДНК должна выступить в роли промотора, который укажет клетке на необходимость создания РНК-копий из остальных фрагментов. Во-вторых, эти копии РНК должны обладать последовательностью, которую можно будет преобразовать в схему жизнеспособной матричной РНК для белковой фабрики.
Более того, эта матричная РНК должна закодировать достаточно длинную белковую цепочку (в среднем длина белка составляет 300 аминокислот). Данный вариант крайне маловероятен, поскольку на случайном отрезке ДНК примерно 1 из 20 кодонов окажется «стоп-кодоном». И, наконец, новый белок должен выполнять некую полезную функцию. Все эти трудности казались непреодолимыми.
Данная точка зрения изменилась в 2006 году, когда Дэвид Бегун из Калифорнийского университета и Дэвис с коллегами обнаружили у дрозофил несколько новых генов с последовательностями, не похожими ни на один из старых генов. Они предположили, что эти гены, кодирующие относительно небольшие белки, эволюционировали из «мусорной» ДНК в течение последних нескольких миллионов лет. Пару лет спустя в процессе поиска новых генов у дрозофил были обнаружены еще девять генов, которые, похоже, самостоятельно образовались из «мусорной» ДНК. Другое исследование показало, что с тех пор, как ветви эволюции человека и шимпанзе разошлись более 6 миллионов лет назад, из некодирующей ДНК появилось как минимум шесть новых человеческих генов.
Чем же объяснить столь большую цифру при ничтожно малой вероятности самостоятельного образования гена? Частичным ответом может служить недавнее открытие: несмотря на то что половина нашего генома является «мусорной», 90 % генов можно случайно транскрибировать в РНК.
Это означает, что случайные фрагменты «мусорной» ДНК могут превращаться в белок не так уж и редко. Поскольку, скорее всего, большая часть случайных белков окажется вредной, естественный отбор уничтожит эти последовательности ДНК. Однако время от времени возникает одна удачная мутация. Последовательность, которая делает что-то полезное, будет передаваться внутри популяции и быстро превратится в новый ген, оптимизированный под любую нужную роль.
Пройдет еще много лет, прежде чем мы до конца поймем важность различных механизмов образования новых генов. Однако уже сейчас очевидно, что классический взгляд на эволюцию генов не дает нам полного объяснения. Эволюция не любит суету: она берет новые гены везде, где ей это удается.







