Полная версия
Кости, гены и культура



Более того, даже если ранние гоминиды не охотились на крупную дичь и не вступали в открытую конфронтацию с хищниками, они могли потихоньку воровать добычу у каких-нибудь зазевавшихся саблезубых кошек. Это тоже могло негативно сказаться на долгосрочных перспективах выживания хищников.
Понемногу накапливаются факты, согласующиеся с гипотезой о влиянии древних гоминид на вымирание африканских хищников (Werdelin, Lewis, 2013). В 2020 году шведские палеонтологи получили новые аргументы в ее пользу (Faurby et al., 2020). Работа шведских ученых основана на анализе данных по относительно небольшому, но исключительно хорошо изученному в палеонтологическом отношении району Восточной Африки. Этот район, охватывающий часть территорий Эфиопии, Кении и Танзании, на протяжении последних 4 млн лет был действующей ареной эволюции гоминид (если и не единственным или главным центром). Ископаемая летопись гоминид здесь весьма богата и прослеживается от ардипитеков, живших около 4 млн лет назад, до поздних сапиенсов включительно.
Местная плиоценовая (5,33–2,58 млн лет назад) и плейстоценовая (2,58–0,0117 млн лет назад) фауна хищных млекопитающих изучена очень подробно, причем один из авторов исследования, Ларс Верделин, внес важный вклад в ее изучение. Именно он собрал и выверил массив данных по восточноафриканским ископаемым хищникам, на котором основана работа. То, что все находки проверялись и интерпретировались одним и тем же специалистом, в данном случае хорошо. Это позволило минимизировать неопределенность, часто возникающую в подобных исследованиях из-за того, что у разных палеонтологов могут быть разные взгляды на систематику изучаемых групп и таксономическую принадлежность находок.
Массив включает 88 видов хищных млекопитающих, живших на изучаемой территории в течение последних 4 млн лет, и 279 “нахождений”, то есть комбинаций “место / время / вид животного”. Лишь девять из 88 видов существуют и поныне, все остальные вымерли. Среди вымерших крупных хищников – гигантские гиены, выдры, куницы и циветты, саблезубые кошки и медведи Agriotherium sp. (рис. 1.4).
Всех хищников подразделили на две группы: крупные с массой тела более 21 килограмма и мелкие. Такое разделение является более или менее общепринятым среди специалистов по экологии хищных зверей, поскольку отражает существенные различия в способах добычи пропитания и спектре доступных жертв. Мелкие хищники, как правило, охотятся на тех, кто помельче, затрачивая на каждую охоту не слишком много времени и сил. Крупные хищники полагаются в основном на крупную добычу, поэтому охотятся реже, но с гораздо большими затратами. Эти различия важны в контексте предполагаемого влияния гоминид – например, клептопаразитизм вряд ли способен сильно навредить мелким хищникам, но может представлять серьезную проблему для крупных.
Реконструированная динамика видового разнообразия крупных хищников в рассматриваемом районе показана на рисунке 1.5.
Данные указывают на стабильно высокое разнообразие крупных хищников от 4 до 2 млн лет назад и на последующий быстрый упадок. Примечательно, что упадок происходил почти синхронно во всех группах, за исключением разве что псовых (которые, впрочем, никогда не играли большой роли в местных экосистемах). Это трудно объяснить какими бы то ни было внутренними закономерностями эволюции крупных хищников, не привлекая внешних факторов, таких как климатические изменения или, например, эволюция гоминид.
Ученые сопоставили данные по хищникам с оценками температуры, количества осадков, площади лесов и дефицита влаги в изучаемом районе, а также с данными по объему мозга местных гоминид (рис. 1.6).
Статистический анализ не выявил никакой связи между вымиранием крупных хищников и колебаниями климата (температуры, количества осадков). Из рассмотренных переменных только две могут претендовать на роль факторов, способствовавших упадку крупных хищников, – прогрессивная эволюция гоминид, отразившаяся в увеличении объема мозга, и сокращение площади лесов. Причем первый фактор, по-видимому, влияет сильнее, чем второй (точнее говоря, его связь с вымиранием хищников имеет более высокий уровень статистической значимости).
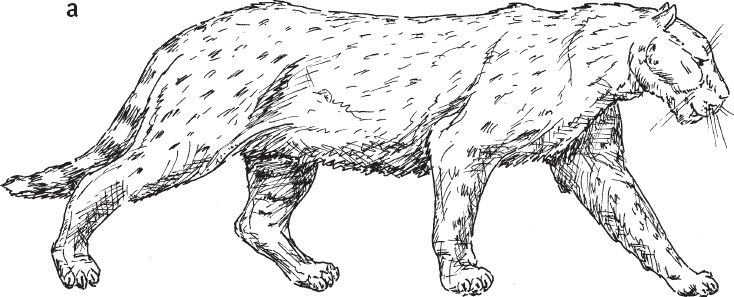
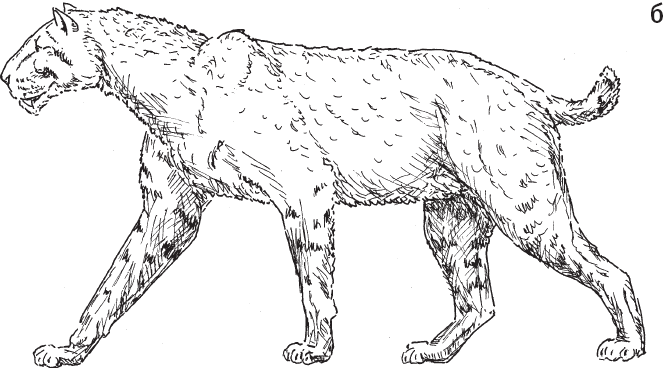
Рис. 1.4. Некоторые крупные хищники, когда-то обитавшие в Восточной Африке вместе с гоминидами. а – саблезубая кошка Dinofelis sp. В изученном районе в период от 4,23 до 4,09 млн лет назад встречалось как минимум три вида этого рода. б – саблезубая кошка Homotherium sp. (обитала от 4,1–4,35 до 1,39–1,64 млн лет назад). в – гигантская гиена Pachycrocuta sp. (от 3,63–3,85 до 3,0–3,2 млн лет назад). г – гигантская выдра Enhydriodon dikikae. В изученном районе водилось четыре вида этого рода, и все были крупные. Возраст самых древних находок гигантских выдр на этой территории – 4,07–4,23 млн лет, самых молодых – 1,88–2 млн лет.
Что касается мелких хищников, то скорость их вымирания в течение последних 4 млн лет оставалась практически постоянной. По-видимому, на их вымирание не оказывали существенного влияния ни климатические изменения, ни прогрессивное развитие гоминид.
Таким образом, результаты показывают, что упадок крупных хищников в Восточной Африке мог быть связан либо с эволюцией гоминид, либо с сокращением лесов. Выделить одну из этих гипотез на основании данных по единственному району не удается, потому что между ростом мозга гоминид и сокращением лесов в Восточной Африке прослеживается сильная корреляция (оба процесса шли параллельно: мозг рос, а леса сокращались).
Чтобы разрешить эту дилемму, ученые использовали данные по соотношению крупных и мелких хищников в разных районах современной Африки. Идея была в том, что те факторы, которые вызвали упадок крупных (но не мелких) хищников в Восточной Африке в плейстоцене, наверное, должны срабатывать и сегодня, снижая долю крупных видов в общем разнообразии хищных млекопитающих.
Выяснилось, что в наши дни доля крупных хищников в разных районах Африки связана обратной зависимостью со степенью антропогенного воздействия и количеством осадков, но практически не зависит от температуры. Влияние площади лесов тоже прослеживается, хотя и слабое. Опираясь на эти современные корреляции между долей крупных хищников и количеством осадков или площадью лесов, исследователи попытались ретроспективно рассчитать динамику доли крупных хищников за последние 4 млн лет (без учета антропогенного фактора). Получилось, что если бы соотношение крупных и мелких хищников контролировалось количеством осадков и площадью лесов так же, как в наши дни, то оно должно было бы оставаться примерно постоянным в плиоцене и плейстоцене. Что явно противоречит наблюдаемому упадку крупных хищников (на фоне сохранения разнообразия мелких) в последние 2 млн лет. Это можно рассматривать как косвенный аргумент в пользу того, что главной причиной упадка крупных хищников были все-таки люди.
Еще один аргумент удалось получить, сравнив африканские данные с аналогичной, хотя и менее детальной и выверенной информацией по плиоцену и плейстоцену Северной Америки. Дело в том, что в течение нескольких последних миллионов лет сокращение лесов происходило не только в Африке. Это был глобальный процесс, захвативший в том числе и Северную Америку. Однако там вплоть до самого конца плейстоцена не было гоминид.
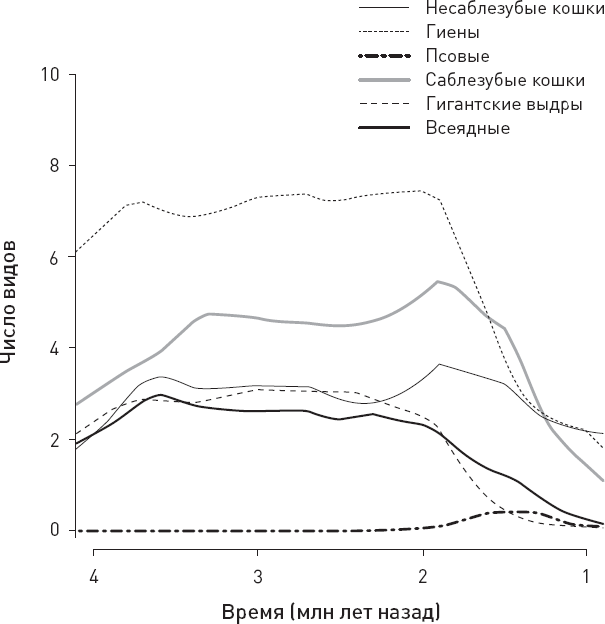
Рис. 1.5. Реконструкция динамики видового разнообразия крупных хищников в рассматриваемом районе. Показаны данные по шести важнейшим группам: кошачьи с коническими зубами, то есть несаблезубые, гиены, псовые (все три группы существуют по сей день), саблезубые кошки, гигантские выдры и “всеядные” (все три исчезли). В последнюю группу входят вымерший медведь Agriotherium, гигантская куница и три вида гигантских циветт. То, что число видов (по вертикальной оси) обычно нецелое, связано с тем, что разнообразие реконструировалось сложными статистическими методами на основе заведомо неполной выборки и неточных датировок. По рисунку из Faurby et al., 2020.
Палеонтологические данные показывают, что сокращение лесов в Северной Америке не сопровождалось упадком крупных хищников. Даже наоборот: доля крупных хищников в общем разнообразии североамериканских хищных млекопитающих увеличилась в плейстоцене по сравнению с плиоценом (с 34–37 до 42–43 %). Это делает еще менее правдоподобной гипотезу, согласно которой упадок восточноафриканских крупных хищников был вызван сокращением лесов.
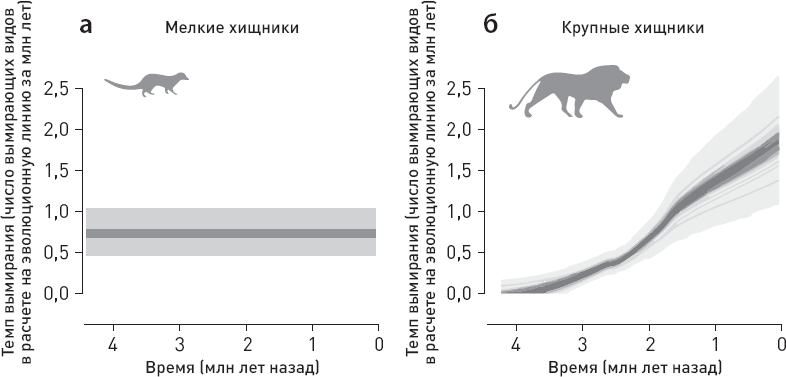
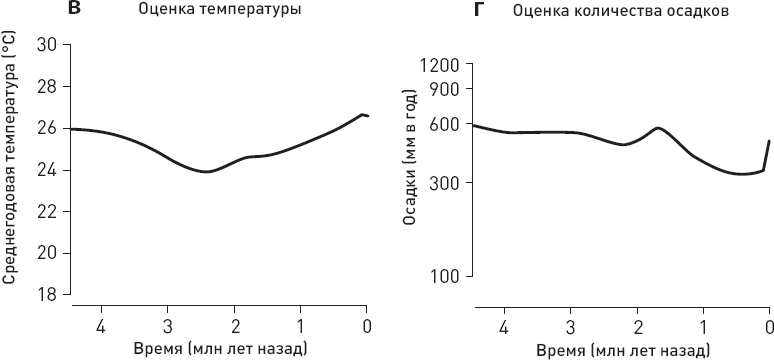
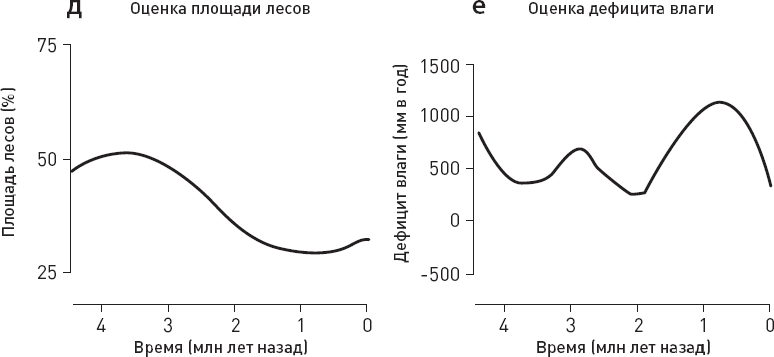
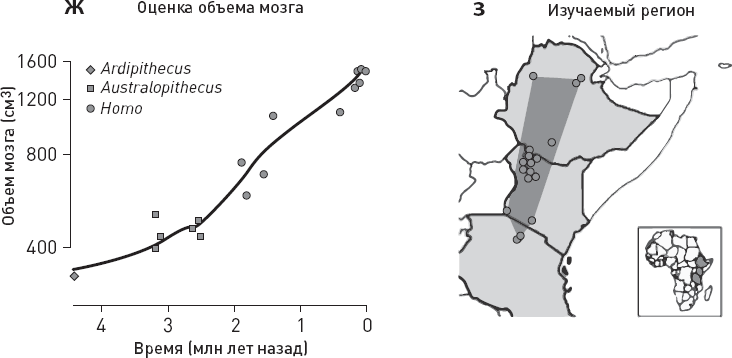
Рис. 1.6. Вымирание восточноафриканских хищников и его возможные причины. а, б – модели, лучше всего описывающие динамику вымирания мелких и крупных хищников. Для мелких хищников (а) оптимальной оказалась модель с постоянной скоростью вымирания видов, для крупных (б) – модель, предполагающая линейную зависимость скорости вымирания хищников от объема мозга местных гоминид. в—е – оценки температуры, количества осадков, площади лесов и дефицита влаги в изучаемом районе (дефицит воды оценивался по соотношению изотопов кислорода в зубной эмали травоядных). ж – оценка объема мозга местных гоминид. з – карта изученного района, кружочками показаны местонахождения ископаемой фауны (палеоклиматические реконструкции основаны на данных из тех же местонахождений). По рисунку из Faurby et al., 2020.
Таким образом, результаты свидетельствуют в пользу важной роли древних гоминид в вымирании крупных хищников в Восточной Африке. Пока трудно сказать, каким был основной механизм влияния гоминид на хищников (охота, клептопаразитизм или что-то еще) и когда это влияние стало набирать силу. Ученые попробовали разобраться в этом, сравнивая модели двух типов: первые предполагали, что скорость вымирания крупных хищников всегда была прямо пропорциональна объему мозга гоминид, а вторые – что гоминиды начали влиять на вымирание хищников не сразу, а лишь начиная с какого-то момента (например, это мог быть момент изобретения охоты на крупных животных, приручение огня или что-то еще). В итоге модели второго типа проиграли в статистическом состязании моделям первого типа. Таким образом, нет оснований считать, что некие примитивные варианты поведения гоминид еще не влияли на вымирание хищников, а более продвинутые стали влиять. Похоже, это влияние началось еще в очень давние времена, когда ни о какой охоте на крупную дичь не могло быть и речи.
Может показаться, что слишком смелые выводы были сделаны на основе пусть и тщательно собранного и детально изученного, но все-таки скудного материала. Напомним, что речь в конечном счете идет всего лишь о 279 палеонтологических находках с надежными привязками к месту и времени. Однако такая ситуация как для палеонтологии, так и для археологии палеолита не только типична, но и неизбежна: материал всегда катастрофически неполон, датировки приблизительны, видовые определения часто спорны. В данном случае исследователи сделали все возможное, чтобы извлечь из имеющегося материала максимум полезной информации.
Глава 2
От хабилисов к эректусам, хоббитам и прочим
Человеческий род, едва утвердившись в Африке, начал расширять свои владения. Люди расселялись по разным регионам, осваивали новые ресурсы, приручали огонь, начинали проявлять первые признаки символического мышления. Разнообразили свою морфологию и культуру, пересекали морские проливы, добирались до уединенных уголков, где порой превращались в обособленные виды. В разных популяциях эволюция шла разными путями, меняя то одни черты, то другие. У кого-то мозг при этом рос (как у наших прямых предков), у кого-то нет, а у кого-то и вовсе уменьшался. Биологическая и культурная эволюция ранне- и среднеплейстоценовых Homo не была похожа на дружное, всеобщее движение к сияющим вершинам разума и цивилизации. Однако постепенное совершенствование орудийной деятельности и мозга в отдельных популяциях позволяло им побеждать в конкуренции с другими животными и друг с другом, понемногу вытеснять группы с менее эффективной культурой, осваивать новые территории и адаптироваться к неблагоприятному климату.
Пищевая революция эректусов: прощание с лесами
Одним из самых информативных источников сведений о диете ископаемых гоминид является изотопный состав углерода в зубной эмали. У углерода два стабильных изотопа: легкий 12С и тяжелый 13C. В природе они встречаются в соотношении, близком к 93:1. Если в зубной эмали у животного повышена доля тяжелого изотопа 13C (ее обозначают δ13Cena, где ena – сокращение от слова enamel, “эмаль”), то это, скорее всего, значит, что животное питалось в основном дарами открытых пространств, поросших травами, – саванн или степей (их еще называют травяными биомами). То есть либо травой, либо травоядными животными. Низкие же значения δ13Cena указывают на питание дарами леса.
Дело в том, что большинство лесных растений использует обычный, “классический” вариант фотосинтеза (так называемый С3-фотосинтез), в ходе которого в качестве сырья для синтеза органики используются преимущественно молекулы CO2 с легким изотопом углерода 12С. Название “C3-фотосинтез” связано с тем, что углерод из углекислого газа включается в состав трехуглеродных органических молекул (а не четырехуглеродных, как при C4-фотосинтезе), хотя для дальнейшего нашего обсуждения это неважно.
Во второй половине кайнозойской эры, начиная примерно с 30 млн лет назад, уровень CO2 в атмосфере стал снижаться, а климат на планете – портиться. В высоких широтах похолодало: сначала покрылась льдом Антарктида, а позже, в плейстоцене, начались периодические оледенения в Северном полушарии. При этом в низких широтах во многих районах климат постепенно становился более засушливым. Это вело к сокращению площади лесов и расширению травяных биомов, в том числе африканских саванн. В новых условиях получили преимущество травы, практикующие другой вариант фотосинтеза – так называемый C4-фотосинтез, при котором в состав органических веществ включается больше тяжелого изотопа 13С. Растения с С4-фотосинтезом лучше приспособлены к сухому жаркому климату в сочетании с нехваткой CO2. Вот поэтому мы и можем определить, чем питалось то или иное африканское животное: если в эмали его зубов много 13C, то дарами саванны, если мало – дарами леса.
Но вернемся к гоминидам. Изучение изотопного состава углерода в зубной эмали ископаемых гоминид показало, что 4 млн лет назад ранние австралопитеки, такие как анамский австралопитек (Australopithecus anamensis), получали углерод в основном из лесных (C3) растений. У жившего 3,3 млн лет назад кениантропа Kenyanthropus platyops диета была более разнообразной, судя по разбросу значений δ13Cena у разных индивидов.
Около 2 млн лет назад восточноафриканские гоминиды подразделились на две ветви. Одна из них (ранние Homo) поначалу питалась C3– и C4-ресурсами с некоторым предпочтением (65 %) лесной продукции, а другая (парантропы) отдавала должное продукции саванн (75 %).
Позднее, около 1,7–1,5 млн лет назад, у людей, которые к тому моменту уже были представлены более продвинутым видом – человеком прямоходящим (Homo erectus), произошел сдвиг в сторону потребления ресурсов саванн (C4). В итоге люди сравнялись в этом отношении с парантропами, у которых все осталось по-прежнему (Cerling et al., 2013).
Антропологи давно склоняются к мысли, что лейтмотивом ранней эволюции рода Homo было освоение саванны и новых способов добычи пропитания – вплоть до полноценной охоты на крупных травоядных. Но датировки остаются спорными. Некоторые специалисты считают, что самые радикальные перемены должны были произойти не 1,7–1,5 млн лет назад, а свыше 2 млн лет назад, при переходе от поздних австралопитеков к первым Homo. Ведь в это время начинается увеличение мозга и систематическое использование каменных орудий для разделки туш.
Впрочем, идея о радикальном переломе 1,7–1,5 млн лет назад тоже выглядит логичной, потому что примерно в это время ранних Homo – человека умелого (H. habilis) и человека рудольфского (H. rudolfensis) – с их олдувайской культурой, крупными челюстями и увеличенными молярами сменяют рослые H. erectus с более совершенной ашельской культурой и менее мощным жевательным аппаратом.
Так почему же изменился рацион ранних гоминид? Нужно ли усматривать в этом признаки активно идущей биологической и культурной эволюции, затрагивающей пищевое поведение, или же это просто пассивный ответ на изменения условий среды? В первом случае рацион должен был измениться без всяких климатических причин, в силу развития мозга и культурного прогресса, позволившего расширить спектр доступных ресурсов. А во втором ведущей силой должна была стать смена окружающей обстановки: саванны расширялись, леса сокращались, а люди, как могли, подстраивались под новые условия. Может быть, люди и не меняли принципиально своего пищевого поведения, а продолжали использовать те же C3– и C4-ресурсы, что и раньше, просто вокруг стало больше вторых.
Чтобы разобраться в этом, нужно понять палеоэкологический контекст. Прежде всего выяснить, как менялись растительность и диета млекопитающих в тех местах, где обитали гоминиды. Если у других животных все происходило так же, как у гоминид, то, по-видимому, следует принять версию о пассивном ответе гоминид на смену ландшафтов. Если иначе, то правдоподобнее будет выглядеть гипотеза об активном освоении гоминидами новых ресурсов.
В 2019 году большой коллектив американских палеоантропологов и геологов опубликовал результаты масштабного исследования, посвященного выяснению причин увеличения δ13Cena у людей, живших в период от 2 до 1,4 млн лет назад (Patterson et al., 2019). Работа базировалась на хорошем ископаемом материале из Кении, из восточных окрестностей озера Туркана. В материале были представлены и растительный, и животный комплекс, и, конечно, гоминиды. Чтобы получить общее представление о смене рациона у млекопитающих, ученые проанализировали соотношение изотопов углерода в 999 образцах зубной эмали. Кроме того, был измерен изотопный состав углерода палеопочв из тех же мест и слоев.
Помимо углерода оценивалось также соотношение изотопов кислорода (δ18O), по которому можно судить о круговороте воды (известно, что вода с тяжелым изотопом 18O испаряется медленнее). У животных с невысоким уровнем испарения с поверхности тела, например у гиппопотамов, носорогов и слонов, значения δ18O мало связаны с засушливостью или влажностью климата и примерно соответствуют содержанию 18O в дождевой воде. И напротив, у животных с высоким уровнем испарения – жирафов и буйволов – δ18O повышается при аридизации климата.
Что же выяснилось? Ни у каких групп млекопитающих, кроме гоминид, не было тенденции к смене рациона с лесного на степной, то есть не было сдвига в сторону С4. Даже у животных со смешанным питанием, которые одинаково охотно употребляют C3– и C4-растения (таковы некоторые полорогие), никакого существенного сдвига в сторону C4 не произошло. А ведь эта группа, по идее, должна была бы в первую очередь изменить спектр питания, если бы экологическая обстановка того требовала. Зато у представителей Homo переход с лесных ресурсов на саванные прекрасно заметен (рис. 2.1). Если сначала (2,0–1,7 млн лет назад) они потребляли больше C3-ресурсов, то затем переориентировались на С4-ресурсы, став в этом плане ближе к парантропам (см. рис. 2.1, б).
Что касается показателя δ18O, то у Homo регистрируется достоверное его снижение, тогда как у парантропов он практически не менялся. Скорее всего, это говорит об изменениях экологии и пищевого поведения людей. Тем более что доля 18O в дождевой воде, по-видимому, не снижалась, а росла. Об этом можно судить по увеличению δ18O в зубной эмали гиппопотамов. Еще нужно учитывать, что низкие значения δ18O в целом более характерны для хищников, чем для растительноядных. Поэтому снижение δ18O у эректусов по сравнению с хабилисами согласуется с идеей о том, что эректусы стали есть больше мяса.
По имеющимся данным можно понять, когда в окрестностях озера Туркана начала меняться растительность. В этом помог анализ изотопов углерода в палеопочвах. По-видимому, саванны с преобладанием C4-растительности распространились в этом районе гораздо раньше 2 млн лет назад. А между 2 и 1,4 млн лет назад если и происходило какое-то дополнительное расширение саванн, то очень незначительное. Вспомним данные о снижении площади лесов в Африке, приведенные выше (см. рис. 1.6): леса начали заметно сокращаться около 3 млн лет назад. Это согласуется с тем, что численность травоядных, потребителей C4-растений, по сравнению с другими группами 2,0–1,4 млн лет назад не увеличивалась. Из этого опять-таки следует, что рост доли C4-ресурсов в диете эректусов по сравнению с хабилисами не может быть пассивным следствием изменения доступной им растительности (и животных, которые ей питались). Ведь растительность почти не менялась. Менялся образ жизни людей и стратегии добычи пропитания.
Все это вместе – серьезный аргумент в пользу того, что изменения, происходившие у Homo, не были пассивным ответом на изменения среды. По-видимому, они были связаны именно с эволюцией людей – биологической или культурной (хотя противопоставлять эти два процесса у Homo вряд ли правильно: скорее всего, они были с самого начала сплетены в неразрывное целое).
Таким образом, получается, что 1,7–1,5 млн лет назад в диете представителей рода Homo – а это уже первые эректусы – произошли серьезные изменения, не связанные с изменениями окружающей растительности и ландшафтов и не наблюдавшиеся у других млекопитающих, включая приматов (парантропов и ископаемых павианов). Эректусы стали потреблять больше ресурсов саванны (C4) по сравнению с ранними Homo.
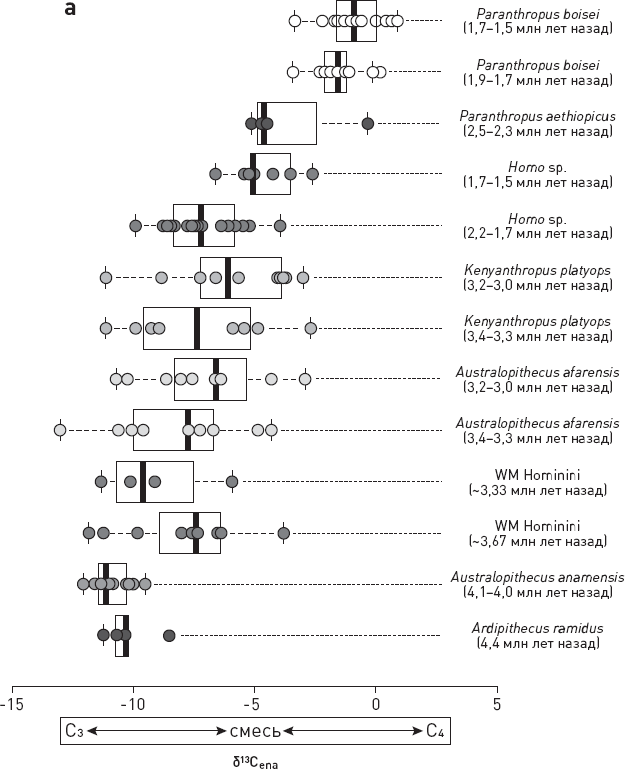
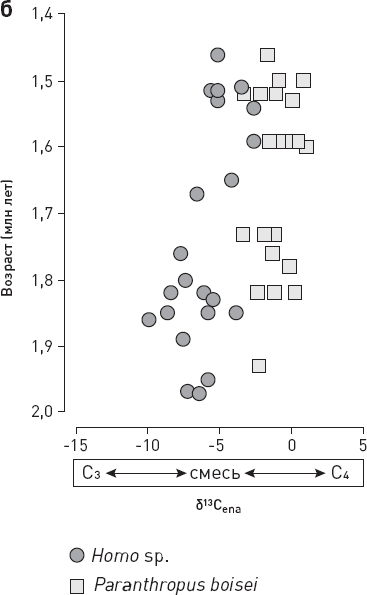
Рис. 2.1. Уточненные данные по изотопному составу углерода в зубной эмали (δ13Cena) у восточноафриканских гоминид. Чем правее расположена точка, тем выше в диете доля ресурсов саванны (C4). WM Hominini – австралопитеки из Ворансо-Милле. Отдельно (б) показаны подробные данные по людям и парантропам с указанием возраста каждого образца (по вертикальной оси). Видно, что парантропы были и оставались специализированными потребителями ресурсов саванны, а вот люди постепенно меняли свои предпочтения, приблизившись к парантропам после отметки в 1,7 млн лет назад. По рисунку из Patterson et al., 2019.
Конец ознакомительного фрагмента.
Текст предоставлен ООО «ЛитРес».
Прочитайте эту книгу целиком, купив полную легальную версию на ЛитРес.
Безопасно оплатить книгу можно банковской картой Visa, MasterCard, Maestro, со счета мобильного телефона, с платежного терминала, в салоне МТС или Связной, через PayPal, WebMoney, Яндекс.Деньги, QIWI Кошелек, бонусными картами или другим удобным Вам способом.