


Полная версия
Культурно-историческая психология волевого действия: От прогноза – к поступку
Рисунок 4а (опыты серии А) показывает, что до конца опыта не угасает депрессия основного ритма на ЭЭГ сенсомоторных областей обоих полушарий. Число депрессий на ЭЭГ затылочной области в этой серии постепенно снижается. Угасание КГР на световые стимулы выражено неотчетливо.
На рисунке 4б (опыты серии В) видно, что депрессия основного ритма на ЭЭГ сенсомоторной области полушария, контрлатерального работающей руке, практически не угасает до конца опыта. Реже наблюдается депрессия на ЭЭГ сенсомоторной области другого полушария и на ЭЭГ затылочной области. КГР в конце опыта этой серии наблюдается очень редко.
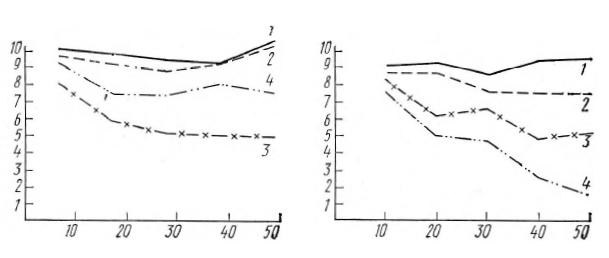
Рис. 4. Изменение числа реакций депрессии основного ритма ЭЭГ и числа КГР на 10 сигналов в ходе опыта в среднем по всем испытуемым: а (слева) – для сигналов с Р2 = Р2=0,5, б (справа) – для сигналов Р1 = 1; Р2=0.
Обозначения: 1 – сенсомоторная область полушария, контрлатерального «работающей» руке; 2 – сенсомоторная область полушария, контрлатерального «неработающей» руке; 3 – затылочная область; 4 – число КГР по десяткам сигналов. По вертикали отложено число реакций на 10 сигналов, по
Когда после звука предъявляется один и тот же световой сигнал, то на ЭМГ работающей руки (той руки, которой испытуемый реагирует на появляющийся световой сигнал) в интервале между выключением звука и предъявлением светового сигнала выявляется повышенная биоэлектрическая активность по сравнению с неработающей рукой. Амплитуда импульсов на ЭМГ работающей руки в интервале между выключением звука и предъявлением светового сигнала в течение опыта меняется у разных испытуемых на величину от 5 до 40 мкв и в среднем за 10 сигналов на 5–7 мкв больше, чем на ЭМГ неработающей руки. Если после звука предъявляются в случайном порядке два световых сигнала с равной вероятностью (серия А), то в течение опыта наблюдается примерно равная по амплитуде биоэлектрическая активность обеих рук. В этой ситуации в среднем за 10 сигналов разность между амплитудой импульсов на ЭМГ правой и левой рук в интервале между выключением звука и предъявлением светового сигнала уменьшается к концу опыта (31-50-й сигналы) до одного – двух микровольт (см. рисунки 5–7).
На рисунке 5 видно, что в серии A (P1 = P2 = 0,5) на девятом предъявлении сигналов депрессия основного ритма на ЭЭГ теменно-затылочной области возникает после выключения звука (сигнал «внимание»); в остальных областях депрессия возникает в начале действия
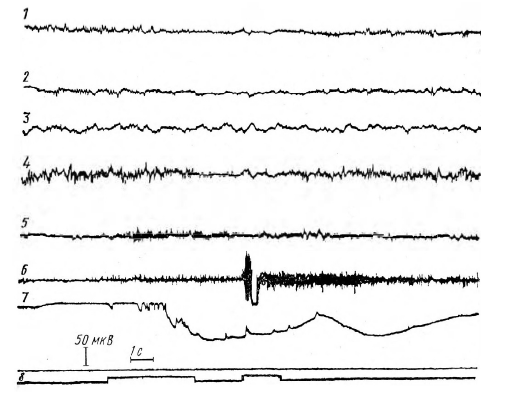
Рис. 5. ЭМГ- и ЭЭГ-реакции испытуемого В. И. на 9-е предъявление сигналов в серии A (P1 = P2=0,5).
Обозначения: 1 – ЭЭГ, затылочная область; 2 – сенсомоторная область левого полушария; 3 – сенсомоторная область правого полушария; 4 – теменно-затылочная область; 5 – ЭМГ левой руки; 6 – ЭМГ правой руки; 7 — КГР; 8 – отметка раздражителей (первый сигнал – предупреждающий звук, второй – пусковой – свет)
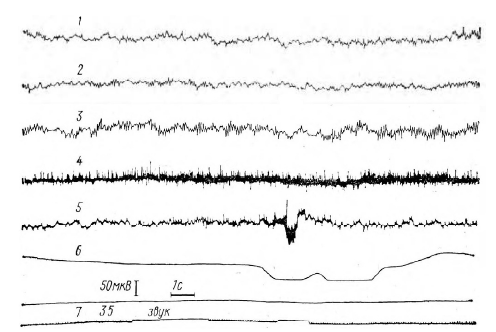
Рис. 6. ЭМГ- и ЭЭГ-реакции испытуемого Б. А. на 35-е предъявление сигналов в серии A.
Обозначения: 1 – ЭЭГ сенсомоторная область левого полушария;
2 – правого полушария; 3 – теменно-затылочной области; 4 – ЭМГ левой руки; 5 – ЭМГ правой руки; 6 – КГР; 7 – отметка сигналов
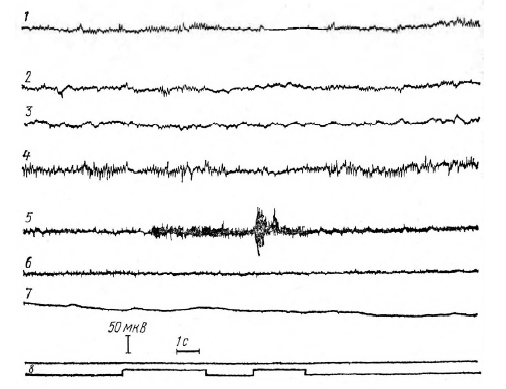
Рис. 7. ЭЭГ- и ЭМГ-реакции испытуемого Л. А. на 29-е предъявление сигналов серии В (P1 = 1, Р2=0). Обозначения те же, что и на рисунке 6 звука. На ЭМГ обеих рук включение сигнала «внимание» ведет к повышению амплитуды импульсов. КГР возникает и на сигнал «внимание» и на пусковой сигнал (свет). На 35-ом предъявлении сигналов (рисунок 6) депрессия на ЭЭГ затылочной и теменно-затылочной областей возникает только при включении светового сигнала, в то время как в сенсомоторных областях она возникает раньше – в интервале между выключением сигнала «внимание» и включением пускового сигнала (свет). Несколько опережает пусковой сигнал и КГР.
На рисунке 7 (опыты серии В; P1= 1, Р2=0) видно, что во время предъявления сигнала «внимание» увеличивается амплитуда импульсов только на ЭМГ работающей руки. Депрессия основного ритма на ЭЭГ сенсомоторных областей возникает раньше, чем на ЭЭГ затылочной и теменно-затылочной областей.
На рисунке 8 показано, что разность между амплитудой импульсов на ЭМГ правой и левой рук уменьшается к концу опыта до 1–2 мкв в серии А и увеличивается в среднем до 6 мкв в серии В в пользу работающей руки. В серии В после первого десятка сигналов все значения разности (кроме одного) расположены выше нуля.
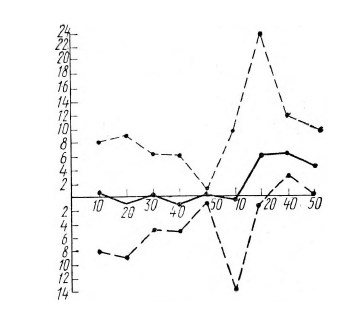
Рис. 8. Изменение в ходе опыта разности между амплитудой ЭМГ (в мкв) правой и левой руки в серии А, работающей и неработающей руки в серии В.
По вертикали отложена величина разности в мкв, по горизонтали – номера сигналов. Приведены величины по десяткам сигналов. Первые 50 сигналов – серия А, вторые 50 – серия В. Разность в мкв в пользу правой (серия А) и «работающей» (серия В) руки отложена выше средней линии, разность в мкв в пользу левой и «неработающей» руки отложена ниже средней линии. Сплошная линия – среднее арифметическое разности по всем испытуемым (с учетом знака разности: выше нуля – плюс, ниже нуля – минус), прерывистая линия – максимальное значение разности по всем испытуемым
Таким образом, если после предупреждающего сигнала «внимание» предъявляются два световых сигнала в случайном порядке и с равной вероятностью (серия А), то длительно не угасает депрессия основного ритма на ЭЭГ сенсомоторных областей обоих полушарий и в интервале времени между сигналом «внимание» и предъявлением светового сигнала (пусковой стимул) на ЭМГ обеих рук наблюдается примерно одинаковая по амплитуде биоэлектрическая активность. Практически в этой ситуации не угасает КГР на световые сигналы. Из регистрируемых в опыте реакций угасает только депрессия основного ритма на ЭЭГ затылочной области. В наших опытах световые сигналы были предельно просты и никаких трудностей для восприятия не представляли, поэтому в данном случае не требовалось дополнительной активации зрительной системы и депрессия в затылочной области постепенно угасла.
Если же после сигнала «внимание» каждый раз предъявляется один и тот же световой сигнал (серия В), то в интервале между выключением сигнала «внимание» и предъявлением светового сигнала сохраняется депрессия на ЭЭГ только в сенсомоторной области полушария, контрлатерального работающей руке, а амплитуда импульсов на ЭМГ работающей руки в указанном интервале в среднем (за 10 сигналов) на 5–7 мкв больше, чем на ЭМГ неработающей руки.
Как можно объяснить эти факты? Ответ на предваряющий сигнал в начале эксперимента следует, согласно широко распространенным представлениям, считать проявлением ориентировочной реакции. В ходе эксперимента ориентировочная реакция смещается во времени (наступает со все большим запаздыванием) и одновременно редуцируется (уменьшается число ее компонентов). Реакцию, возникающую в конце эксперимента серии В (P1 = 1, Р2 = 0), можно рассматривать как выработанный условный рефлекс: в ответ на предваряющий сигнал (звук) наступают изменения в двигательном аппарате соответствующей руки, приуроченные по времени к моменту включения пускового сигнала. Анализ всего хода эксперимента позволяет сделать вывод, что перед нами непрерывный процесс, начало которого можно охарактеризовать как ориентировочную реакцию, а конец – как условную.
Иначе обстоит дело в серии А. Здесь условный рефлекс в его классическом виде (оцениваемом по конечному результату – действию) не может быть выработан, так как предваряющий сигнал не несет сведений о том, какой рукой нужно будет реагировать (P1 = P2 = 0,5). В начале эксперимента серии А предваряющий сигнал вызывает ориентировочную реакцию, которая в ходе эксперимента смещается (отстает от предваряющего сигнала и сдвигается к моменту, непосредственно предшествующему пусковому сигналу) и редуцируется. Однако редукция эта меньше, чем в серии В. Реакцию, наблюдаемую в конце эксперимента серии А, можно рассматривать как смещенную и редуцированную ориентировочную реакцию.
Ситуация А и ситуация В отличаются тем, что предупреждающий сигнал в этих ситуациях несет в себе информацию неодинаковой определенности относительно того, какой из пусковых стимулов появится вслед за предупреждающим сигналом. В ситуации А после предупреждающего сигнала с равной вероятностью может появиться любой из двух пусковых стимулов, требующих разных реакций. В ситуации В после предупреждающего сигнала каждый раз появляется один и тот же пусковой стимул. Различные результаты (по реакциям ЭЭГ и ЭМГ) в конце каждой серии опытов можно объяснить тем, что прогноз о появлении определенного пускового стимула в ситуации В более определенен, чем в ситуации А. Результаты обеих серий показывают, что по мере повторения сигналов остаются лишь те реакции, которые осуществляют подготовку к ожидаемым действиям. К концу опыта преднастройка становится более локальной и по времени опережает на 1–2 с появление пускового стимула.
Последовательность изменений реакции в обеих сериях дает, мы считаем, основание полагать, что ориентировочная и условная реакции являются различными сторонами единого физиологического процесса, а не двумя различными, четко разграниченными реакциями.
Ориентировочная реакция, как и условная, является реакцией на предстоящую ситуацию в соответствии с вероятностным прогнозом в этой ситуации. Но если условная реакция соответствует определенному прогнозу, то ориентировочная реакция соответствует прогнозу некоторой неопределенности предстоящей ситуации.
Индивидуальные особенности вероятностного прогнозирования
Вероятностное прогнозирование опирается на прошлый опыт и, естественно, зависит от индивидуальных особенностей данного организма в данный момент. Эти индивидуальные особенности вероятностного прогнозирования выявились и в описанных выше экспериментах в скорости угасания депрессии основного ритма на ЭЭГ сенсомоторной области полушария, контрлатерального неработающей руке, при повторении сигнала.
По скорости угасания депрессии основного ритма на ЭЭГ сенсомоторной области полушария, контрлатерального неработающей руке, в ситуации, когда после звука каждый раз предъявляется один и тот же световой сигнал, испытуемые разделились на две группы: у одной половины испытуемых депрессия постепенно угасала (на 31— 50-й сигналы в среднем наблюдается 5–6 раз на 10 сигналов), у другой половины испытуемых депрессия не угасала до конца опыта, наблюдаясь 8—10 раз на 10 сигналов. У трех испытуемых из последней группы на ЭМГ неработающей руки в интервале между сигналом «внимание» и появлением светового сигнала на первые 20 сигналов выявлялась повышенная биоэлектрическая активность по сравнению с работающей рукой, и в субъективном отчете эти испытуемые указывали, что они ожидали отсутствующего в этой половине опыта сигнала (рисунок 9).
Мы предположили, что различные скорости угасания депрессии на ЭЭГ сенсомоторной области полушария, контрлатерального неработающей руке, объясняются особенностями перестройки прогноза испытуемых. Если это предположение верно, то можно ожидать, что и в других ситуациях, требующих от испытуемых перестройки прогноза, будут наблюдаться аналогичные результаты.
Для проверки этого предположения была проведена серия опытов, в которых испытуемого просили угадывать, какой из двух знаков будет следующим в последовательности экспериментатора.
Особенности эксперимента не позволяли провести эти опыты на тех же самых испытуемых, которые участвовали в опытах с регистрацией компонентов ориентировочной реакции. В опытах участвовало 17 испытуемых.
Испытуемому сообщалось, что экспериментатором будет реализовываться последовательность сигналов, состоящая из плюсов и минусов (в действительности последовательность экспериментатора всегда состояла из одних плюсов). Испытуемого просили угадывать, какой из двух знаков будет следующим в последовательности экспериментатора. После того как испытуемый устно сообщал об ожидаемом знаке, экспериментатор называл следующий знак в своей последовательности (всегда плюс).
По результатам опыта испытуемые разбились на две группы: половина испытуемых (9 человек) с 10—20-й пробы (в среднем с 14-й пробы) начинала называть только плюс, т. е. тот знак, который постоянно называл экспериментатор. До 10-20-й пробы число плюсов и минусов в ответах испытуемых было примерно одинаковым, у двух испытуемых плюсов было немного больше (на 4–5 знаков). Вторая группа испытуемых полностью переходила на плюс с 22—70-й пробы, в среднем по восьми испытуемым с 39-й пробы. Четверо испытуемых этой группы с 5-й по 30-ю пробу называли большее число минусов (отсутствующий в последовательности экспериментатора знак), причем эти испытуемые назвали подряд 12–19 минусов, тогда как при вероятности сигналов P1 = Р2=0,5 эти испытуемые называли подряд не более 5 одинаковых знаков (рисунок 10).
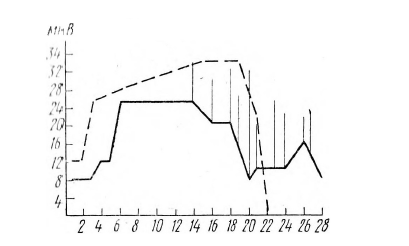
Рис. 9. Изменение амплитуды импульсов на ЭМГ в серии В у испытуемого Л. А.
По горизонтали отложены номера сигналов по порядку, по вертикали – амплитуда импульсов в мкВ. Штриховая линия показывает изменение амплитуды импульсов на ЭМГ правой «неработающей» руки (берется средняя амплитуда за 2 с до предупреждающего сигнала). Сплошная линия – средняя амплитуда импульсов на ЭМГ левой «работающей» руки. Вертикальные линии показывают увеличение амплитуды импульсов на предупреждающий сигнал или в интервале до пускового сигнала
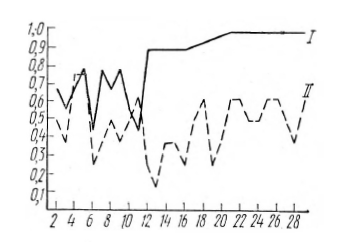
Рис. 10. Перестройка прогноз, а у двух групп испытуемых при неожиданном для испытуемого переходе от последовательности с Р2 = Р2 = 0,5 к последовательности с P1 = 1, Р2=0.
По горизонтали отложены номера ответов испытуемого, по вертикали – частота плюса в ответах испытуемых
Чтобы показать, что деление разных испытуемых на сходные группы (по скорости угасания компонентов ориентировочной реакции и скорости полного перехода на повторяемый экспериментатором сигнал) не является случайным, было проанализировано изменение времени реакции в такой же ситуации у новой группы испытуемых (13 человек). Испытуемых просили нажимать правой рукой на ключ как можно быстрее при появлении цифрового сигнала в виде цифры «2» и левой рукой на левый ключ при появлении цифры «1» (сигналы подавались включением цифровой лампы). Половине испытуемых предъявлялся только сигнал «2», другой половине – только «1». Всего подавалось 100 сигналов. При первых 10 сигналах время реакции у всех испытуемых было на 55–70 % больше, чем время простой двигательной реакции этих испытуемых. По мере предъявления сигналов время реакции у всех 13 испытуемых постепенно уменьшилось, причем это уменьшение нельзя было объяснить тренировкой самого движения. До этого все испытуемые работали с этими же ключами, и время простой двигательной реакции у них не изменялось при длительной подаче сигналов.
По скорости изменения времени реакции испытуемые разделились на две группы (независимо от того, какой рукой они реагировали – правой или левой).
Таблица 2
Снижение времени реакции по ходу исследования у двух групп испытуемых
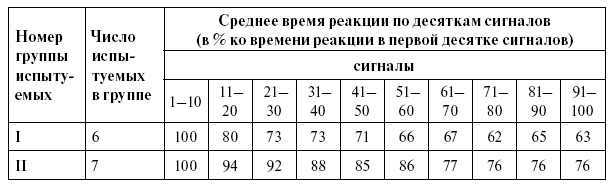
Из таблицы 2 видно, что у одной группы испытуемых (6 человек) время реакции изменяется довольно быстро. При 11-20-м сигналах время реакции у этой группы испытуемых уменьшилось на 20 %, а при 51-60-м сигналах время реакции составляло 66 % от времени реакции на первые 10 сигналов. К концу опыта у испытуемых этой группы время реакции достигало времени простой двигательной реакции или было очень близко к нему. У второй группы испытуемых (7 человек) время реакции изменялось значительно медленнее. При 11-20-м сигналах время реакции уменьшилось в среднем на 6 %, а при 51-60-м сигналах – составило 86 % от времени реакции на первые 10 сигналов. У одного испытуемого из этой группы время реакции на 11-30-й сигналы составляло 103 % по сравнению со временем реакции на первые десять сигналов.
Можно думать, что деление испытуемых на две группы по скорости снижения времени реакции в ходе исследования так же, как и деление их на две группы во скорости угасания депрессии основного ритма на ЭЭГ сенсомоторной области полушария, контрлатерального неработающей руке, и деление испытуемых на две группы по скорости перехода на угадывание знака, который каждый раз повторяется экспериментатором, объясняется особенностями перестройки прогноза испытуемых. Одна группа испытуемых, зная из инструкции и прошлого опыта о том, что должно быть два сигнала, несмотря на то что в опыте каждый раз предъявляется один и тот же сигнал, всё-таки ожидает в соответствии с инструкцией тот сигнал, который отсутствует в последовательности экспериментатора. Ожиданием отсутствующего сигнала и объясняются у этой группы испытуемых длительное неугасание депрессии на ЭЭГ, медленное снижение времени реакции и большое число минусов при угадывании следующего знака в последовательности экспериментатора. Вторая группа испытуемых быстро перестраивает свой прогноз относительно частот сигналов и в соответствии с этим осуществляет преднастроечные реакции. У испытуемых этой группы быстро угасает депрессия основного ритма на ЭЭГ сенсомоторной области полушария, контрлатерального неработающей руке, у них наблюдается быстрый переход па знак, который каждый раз повторяется экспериментатором, и время двигательной реакции у них быстро уменьшается до времени простой двигательной реакции, когда вместо двух сигналов, о которых говорилось в инструкции, в опыте каждый раз предъявляется один и тот же сигнал.
Кожно-гальваническая реакция как отражение преднастройки к движениям
В исследованиях последнего времени кожно-гальваническая реакция (КГР) широко используется как индикатор ориентировочной реакции и как индикатор эмоциональных реакций. В описанных выше опытах КГР выступала как один из компонентов ориентировочной реакции, возникавшей при различной степени нег определенности вероятностного прогноза.
В каком же положении находится КГР по отношению к другим компонентам ориентировочной реакции, отражением чего она является?
Взгляды на природу ориентировочной реакции претерпели за последний период существенную эволюцию – от иллюзорной простоты и ясности к четкому пониманию большой сложности и неясности ее механизма. На смену представлению об ориентировочной реакции как о безусловном рефлексе, осуществляемом той или иной рефлекторной дугой, пришли значительно более сложные представления, включающие в структуру ориентировочной реакции такие понятия, как акцептор действия (П. К. Анохин), аппарат сличения (Н. А. Бернштейн), нервная модель стимула (Е. Н. Соколов).
Вопрос о природе ориентировочной реакции тесно связан с вопросом о том, отражением чего являются различные ее компоненты, в частности КГР. Известно, что чем сложнее выбор возможных двигательных реакций, тем медленнее угасает КГР. Это подтвердилось и в наших опытах: чем больше неопределенность вероятностного прогноза относительно предстоящих двигательных реакций, тем более стойкой является КГР. Единого мнения, объясняющего этот факт, до сих пор не имеется.
Одна из существующих точек зрения заключается в дом, что ориентировочная реакция представляет собой преднастроечную реакцию – подготовку организма к действиям в прогнозируемой ситуации в условиях большей или меньшей неопределенности вероятностного прогноза (Фейгенберг, 1963). С этой точки зрения различные компоненты ориентировочной реакции являются отражением преднастройки (мобилизации) различных физиологических систем. Чем более неопределенным является прогноз, т. е. чем шире круг физиологических систем, действие которых может понадобиться в прогнозируемой ситуации для достижения цели, тем более многокомпонентной является ориентировочная реакция. Так, депрессия альфа-ритма является, можно думать, отражением мобилизации сенсорных систем, а КГР – отражением моторной преднастройки, мобилизации эфферентных систем. Большая или меньшая выраженность различных компонентов ориентировочной реакции соответствует степени выраженности преднастройки различных функциональных систем.
Согласно другой точке зрения (Гращенков, Латаш, 1965), депрессия альфа-ритма рассматривается как результат неожиданности действия раздражителя, как отражение направленного внимания, а КГР – как отражение «принятия решения» выбора определенной реакции из нескольких альтернативных возможностей.
Для экспериментального «разведения» этих двух точек зрения на КГР нами был поставлен следующий эксперимент (Пилипчук, Рутман, Фейгенберг, 1968).
Испытуемым давалась инструкция: после сигнала «внимание» (неяркая вспышка) последует серия из четырех звуков; первые три звука необходимо сравнить; в том случае, если хотя бы два из них отличны друг от друга по высоте, то после четвертого звука следует нажать на ключ указательным пальцем правой руки; если же все три звука будут одинаковыми, нажимать ключ не надо.
Опыты проводились в условиях стандартной процедуры электроэнцефалографического исследования. Регистрировались депрессия доминирующего ритма на ЭЭГ и КГР по Тарханову. Звуковыми сигналами служили громкие тоны от звукового генератора (от 600 до 1000 Гц); звуковые раздражители подбирались таким образом, что не вызывали трудности различения. Каждая серия раздражителей состояла из пяти сигналов – вспышка («внимание») и четыре звуковых тона, которые обозначим цифрами I, II, III и IV. Интервалы между сигналами внутри серии варьировались от 5 до 15 с и около 20 с между сериями. В некоторых случаях четвертый звук отличался от предыдущих, что не повлияло на характер реакций испытуемого.
В случайном порядке предъявлялись серии трех типов (в одном и том же опыте): серия А – все звуки были одинаковыми, серия Б – звук II отличался от звука I, и серия В – звук II был таким же, как звук I, а звук III отличался от них.
Идея эксперимента заключалась в следующем. Если КГР отражает момент принятия решения, т. е. выбора какой-либо одной возможности из нескольких альтернативных возможностей, то КГР должна быть особенно отчетливой и стойкой в серии Б после звука II (именно в этот момент снимается неопределенность действий); в сериях же А и В КГР должна быть более выраженной после звука III, так как решение о том, нужно ли нажимать после IV звука, не может быть еще принято после звука II, а принимается после звука III; в серии А принимается решение, что не нужно нажимать, в серии В – решение, что нужно нажимать.
Если же КГР является отражением преднастройки к действию (движению), то нужно ждать наиболее выраженной КГР в сериях Б и В в интервале между звуками III и IV независимо от того, отличался II или III звук от I звука (КГР приурочена к подготовке действия, а не к моменту принятия решения о том, нужно ли оно). В серий же А КГР после звука III должна быть выражена незначительно, так как после III звука принимается решение, что моторной реакции не требуется и, следовательно, не нужна преднастройка к ней.
Для наглядности изобразим ход эксперимента и ожидаемые результаты в виде схемы:
Серия А: «внимание» – звуки I–II–IIIp – IV
Серия Б: «внимание» – звуки I–IIp – IIIn – IV.
Серия В: «внимание» – звуки I–II–IIInp – IV.
В серии А решение о ненужности моторной реакции принимается после сигнала III, идентичного сигналам I и II. Буквой «р» обозначены моменты времени, в которые нужно ожидать наиболее выраженной КГР, исходя из гипотезы, что КГР отражает момент принятия решения. Буквой «п» обозначены моменты, в которые нужно ожидать наиболее выраженной КГР, исходя из гипотезы, что КГР отражает преднастройку к действию.

