


Полная версия
Избранные труды. Том 1
: В Индии жил великий мыслитель Шри Ауробиндо, в работах которого говорится о «разуме клеток». И.А.Карышева
: Шри Ауробиндо – это не наука, а метафизика. Но индийский биофизик Джагадиш Чандра Бос имел сходные представления. Б. Г. Режабек
В беседе с С. В. Багоцким мы пришли к выводу о том, что МОИП мог бы представить нашу работу на соискание Нобелевской премии. Я делал доклад в МОИП о Нобелевских премиях, не выданных русским ученым, и говорил о том, что в ряде случаев (А. М. Оловников, Ю. А. Лабас) это произошло из-за того, что Европа мало читает нашу научную литературу, а мы не думаем о наших приоритетах и не представляем работ наших учёных в Нобелевский комитет.
: Экспериментальные данные докладчика очень интересны, но объяснение без дальнейших экспериментов – это метафизика. Нужно искать гранты на продолжение работы. Кто хочет – ищет возможности, кто не хочет – ищет причины. У эритроцитов есть метаболическая память. А.Ф.Топунов
Нужны серьезные исследования. Вопрос о выдвижении на Нобелевскую премию наше собрание решать не может, но можно поставить его на общей Конференции МОИП.
: Ваши эксперименты очень интересны и нуждаются в развитии. Но я не могу одобрить Ваше стремление всюду искать душу. Душа – это не объяснение. Е. Д. Яхнин
Председательствующий поблагодарил докладчиков и участников заседания за интересную дискуссию и объявил заседание закрытым.
Председатель секции Проблем
Эволюции материи МОИП Е.Д.Яхнин
Ученый секретарь секции А. К. Погорелов
Ученый секретарь МОИП С. В. Багоцкий
ЗАХВАТЫВАНИЕ РИТМА ПРИ РАЗДРАЖЕНИИ
НЕЙРОРЕЦЕПТОРНОЙ КЛЕТКИ РЕЧНОГО РАКА
БИОФИЗИКА Том XVIII, вып. 21973
А. Б. Коган, Б. Г. Режабек, О. С. Васильева
Ростовский государственный университет
При локальном раздражении синусоидальным током через микроэлектрод быстро адаптирующегося и медленно адаптирующегося нейронов обнаружено захватывание частот с отношением 1: 1 в области 2—4 и 19— 23 гц, а также с отношением 2:1 в области 5—7 и 24 – 27 гц. Увеличение амплитуды стимула приводит к расширению полос захвата. Обсуждается связь наблюдаемых эффектов с проблемой самоорганизации нейронных сетей.
Проблема самоорганизации нейронных структур требует выяснения двух основных вопросов:
1) какие свойства элементов систем существенны для самоорганизации, 2) какими должны быть законы взаимодействия элементов, позволяющие реализовать самоорганизацию.
То, что элементы самоорганизующихся систем должны обладать определенной спецификой, неоднократно подчеркивали как математики [1—3], так и биологи [4—8]. Поведение элементов биосистем в свою очередь определяется спецификой физико-химического состояния живого вещества, находящегося в состоянии «устойчивого неравновесия» [6], которое, с современной точки зрения, можно интерпретировать, как результат взаимодействия многоуровневых внутри- клеточных нелинейных осцилляторов как кинетической, так и молекулярно-структурной природы.
По-видимому, механизм взаимодействия нелинейных осцилляторов является одним из механизмов самоорганизации [9, 10].
В особенности существенным этот механизм может считаться при рассмотрении внутриклеточной самоорганизации как механизма синхронизации пространственно разобщенных процессов в клетке и взаимодействия нейронов, каждый из которых непрерывно подвергается воздействию ритмических импульсных потоков.
Хотя этот вопрос несомненно важен, в современных моделях нейронов, к сожалению, не представлены ни способность нейрона к целесообразному приспособлению, ни возможность наличия эндогенных ритмов, свойст- венных данному нейрону и позволяющих ему избирательно реагировать на входной импульсный поток. Наличие таких ритмов обсуждалось в ряде физиологических работ в связи спроблемой «пейсмекеров» [11].
В настоящей работе представлены эксперименталь- ные данные, свидетельствующие о наличии таких ритмов в изолированной нейрорецепторной клетке речного рака.
Методика
Потенциалы действия (ПД) отводились внеклеточно от нервного ствола с помощью серебряного электрода диаметром 0,3 Импульсы усиливались УБП1—02 и регистрировались с помощью шлейфного осциллографа Н-102. мм.
Раздражение производилось стеклянными микроэлек- тродами, заполненными 3раствором КСl. Диаметр кончика измерялся под микроскопом МБР-1 и составлял 2—5мк, сопротивление 1—5 Мом. М
Микроэлектрод подводили с помощью микроманипулятора ММ-1 к избранной точке нейрона под контролем микроскопа МБР-1 (рис.1). В качестве источника раздражающего тока использовался генератор Г3—47.
Результаты экспериментов
– Реакция быстроадаптирующегося нейрона.
Реакция быстроадаптирующегося (БА) нейрона на механичеcкое (адекватное) раздражение достаточно подробно изучена многими авторами [12, 13].
При электрическом раздражении наиболее эффективным является положение микроэлектрода в области аксонного холмика (АХ), при этом возбуждение происходит, когда на микроэлектроде отрицательное напря- жение. Порог раздражения в этом случае минимален. Располагая микроэлектрод вблизи сомы или удаляясь от АХ по волокну, можно наблюдать изменение порога и даже инверсию знака раздражающего потенциала.
При условии одинаковой реакции на толчок постоянного тока, которой можно добиться, расположив микроэлектрод при подготовке к опыту соответствующим образом, реакции различных препаратов на ритмическое раздражение совпадают с точностью до 10%.
Такое раздражение не вызывает импульсной активности нейрона, пока f c <1,5 гц.
Рис.2
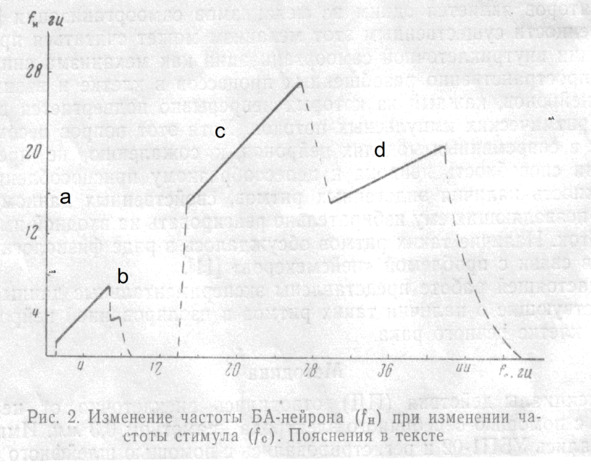
На рис. 2 показано изменение частоты БА-нейрона (f) при изменении частоты стимула (f); амплитуда раздражения 11 в. Микроэлектрод расположен вблизи АХ. H c
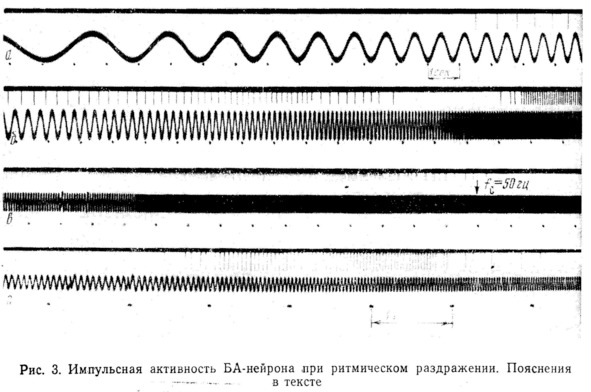
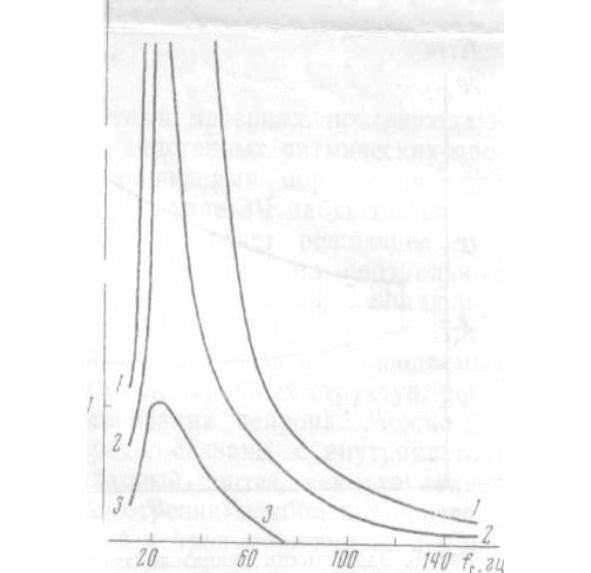
При достижении этой частоты наблюдается резкий переход к режиму, при котором нейрон работает с частотой, совпадающей с f (рис. 3,а). Диапазон захватывания частоты (ЗЧ) зависит от амплитуды стимула. c
В данном опыте ЗЧ скачком прекращается при fc = 7 При изменении f от 7 до 8 наблюдается ЗЧ с делением в отношении 2:1, при частотах, незначительно превышающих 8 нейрон реагирует на каждый третий или четвертый стимул и затем реакция вообще исчезает вплоть до частоты 13 (рис. 3,б). От 13 до 14 реакция является вероятностной, т. е. однозначного соответствия между f и f установить не удается и при повторных стимуляциях реакция в этом диапазоне оказывается каждый раз иной. С 14 начинается ЗЧ вплоть до 27 В диапазоне от 27 до 30 наблюдается переходной режим реакции, при котором можно обнаружить отклик нейрона, соответствующий либо каждому периоду стимула, либо с выпадением одного периода. С 30 до 42 наблюдается ЗЧ с делением частоты 2:1, и, наконец, при более высоких частотах реакция вновь становится вероятностной. гц. гц гц, гц гц гц гц. гц гц с с H
При дальнейшем увеличении частоты стимуляции вплоть до 20 000 импульсная реакция нейрона отсутствует. На рис. 3, представлен ход процесса от 15 до 60 Если изменять амплитуду стимула, общий ход реакции закономерно изменяется. гц в гц.
Результаты можно объединить с помощью рис.4. Линия представляет собой геометрическое место точек, соответствующих началу первой полосы ЗЧ с отношением частот f: fa равным 1:1. Линия б соответ ствует переходу в вероятностный режим, – началу первой полосы ЗЧ с отношением частот 2:1, – ее окончанию и – исчезновению импульсной реакции. Линия соответствует началу новой полосы ЗЧ. а в г д е c
Реакция становится детерминированной (отношение частот 1:1), начиная с линии и остается таковой вплоть до линии Между и реакция недетерминированна, между и захватывание происходит с делением частоты 2:1. В области между и реакция опять становится вероятностной, и, наконец, линия соответствует оконча- тельному исчезновению импульсной реакции нейрона вплоть до f= 20 000 Таким образом, можно наб- людать три четко выраженных полосы ЗЧ и одну ( – менее выраженную. ж, з. з и и к к л л гц. (а-б, ж – з, и -к) в г) с
Рис.4 Пояснения в тексте
Эти эффекты мы наблюдали на 30 препаратах. На одном и том же препарате измерения можно проводить многократно (10 и более раз) без заметных изменений в результатах. Для различных препаратов соответствующие частоты несколько отличаются вследствие небольших различий в положении микроэлектрода, а также в зависимости от физиологического состояния препарата.
Наиболее стабильна область второго ЗЧ с отноше- нием частот 1:1в то время как предыдущие области захватывания на некоторых препаратах отсутствовали. Иногда отсутствовала и область (рис.3, Если кончик микроэлектрода расположить вдали от АХ, общий ход кривой f= φ (f) изменяется. (ж-з), и-к г). н с
В области низких частот в этих опытах до того, как начинается захватывание 1:1, можно наблюдать пачечную активность. Число импульсов в пачке с увеличением f уменьшается, и пачка вырождается в одиночные импульсы, затем происходит деление частоты в 2, 3, … раз и где-то на 10—15 импульсная активность исчезает. Приведенные результаты относятся к опытам, в которых частота стимула линейно нарастала со временем. c п гц
В следующей серии опытов исследовалась реакция БА-нейрона на включение стимула фиксированной частоты (f). c
Рис. 5. Длительность реакции БА-нейрона при включении стимулов различной частоты
Если f находится за пределами зон ЗЧ, то БА-нейрон некоторое время τ, зависящее от fи амплитуды стимула, работает с частотой f= f, затем адаптируется и прекращает импульсную активность (рис. 5). c c н c
При f, принадлежащей области ЗЧ τ = ∞ в этих условиях нейрон не проявляет адаптации. c
Зависимости τ от f для одного из препаратов (два первых диапазона не показаны) представлены на рис. 6. c
Рис.6.Зависимость времени реакции от частоты и амплитуды стимула: 1—амплитуда 20 В; 2 – амплитуда 10 В; 3 – амплитуда 8 В
От препарата к препарату эффект варьирует незначительно.
– Реакция медленно адаптирующегося нейрона.
Реакция МА-нейрона качественно во многом сходна с реакцией БА. Основное отличие связано с тем, что МА-нейрон при отсутствии раздражения может генерировать импульсы с собственной фоновой частотой (2—30 определяемой степенью приложенного растяжения. гц),
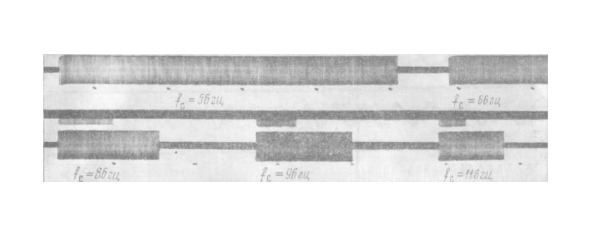
При включении ритмического раздражения, начиная от 0,1частота нейрона модулируется изменениями стимула (рис.7,затем появляются пачки импульсов (рис.7,б и в), дальнейшее увеличение f вызывает ЗЧ с отношением 1:1 (рис.7, гц, а), г). с
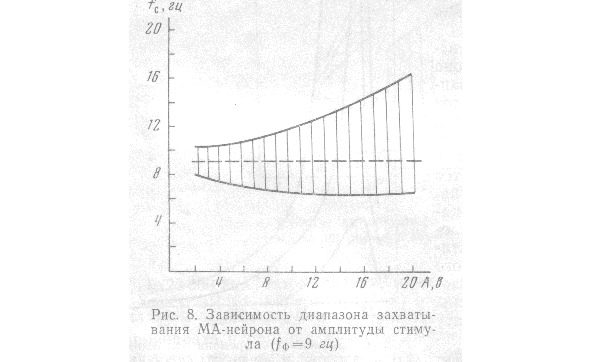
Ширина этого диапазона ЗЧ и значения его граничных частот зависят как от фоновой частоты работы нейрона f, так и от амплитуды стимула. С увеличением fдиапазон захватывания расширяется. Зависимость этого диапазона от амплитуды стимула при f = 9 представлена на рис. 8. ф ф ф гц
На рис. 9 представлены результаты опыта при f = 2 и амплитуде стимула 8ВМикроэлектрод расположен вблизи АХ. ф гц .
Первая область ЗЧ с отношением 1:1 наблюдается при f =1,5—3,5 Затем следует область захватывания 2:1 и довольно широкая полоса вероятностной реакции. с гц.
Интересно отметить, что в области 11
Вхождение в эту область предваряется участком вероятностной реакции, которая может быть обнаружена и при переходе от режима 1:1 к 2:1 и затем после последнего (на рис.9. в интервалах 23—24 и 26—32 соответственно). При частотах выше 32 не удается обнаружить воздействие стимула данной амплитуды на импульсную активность МА, нейрон возвращается к исходной фоновой активности. гц гц
В пяти опытах на МА-ней роне наблюдалась третья область ЗЧ с отношением 2:1 на частотах 40—50 гц.
При большой f и определенной амплитуде стимула обе области ЗЧ с отношением 1:1 сливаются (рис. 7, и ф г д).
При удалении от АХ наблюдается то же изменение эффекта, что и на БА, только при f 10—15 частота нейрона не равна нулю, а соответствует степени растяжения. с> гц
Обсуждение
Полученные результаты дают конкретный материал, подтверждающий представление о наличии в нейроне эндогенных ритмических процессов, связанных с механизмами, управляющими порогом нейрона и генерацией импульсов. То, что в данном случае ЗЧ наблюдается при раздражении изолированного нейрона, представляет решающее преимущество в сравнении с опытами такого же типа на центральных нейронах. Относительно физико-химической природы интрацел- люлярных осцилляторов мы можем высказываться лишь спекулятивно. Представляется, например, привлекате- льной мысль о связи наблюдаемых нами полос захватывания с осцилляциями мембранных структур, регулирующими критический уровень деполяризации нейрона. Можно допустить, что эти структуры, в свою очередь, связаны с внутриклеточными структурами, определяющими белковый синтез, как это делает Хиден [13].
Для кибернетического рассмотрения наибольший интерес имеет представление о том, что отдельный нейрон может обладать набором резонансных ритмов, позволяющих избирательно реагировать на входной поток импульсов. Полученные данные свидетельствуют в пользу такого предположения.
Способность нейрона избирательно реагировать на различные входные частоты необходимо учитывать при разработке моделей нейронов и нейронных сетей. Наличие собственных частот у нейронов может привести к появлению в сетях из таких элементов существенно новых свойств, в частности самопроизвольного распада такой сети на функциональные ансамбли, самоподстройку нейронной сети к наиболее эффективному режиму функционирования, самовосстановлению функциональных свойств при выходе из строя отдельных элементов.



ЛИТЕРАТУРА
1. Бир Ст., Кибернетика и управление производством, «Мир», М., 1966.
2. Цитлин М. Л. Исследования по теории автоматов и моделированию биологических систем, «Наука» М.,1969.
3. Гельфанд И. М., Цетлин М. П., В сб.: Модели структурно-функциональной организации некоторых биологических систем, с. 9,«Наука», М., 1966.
5. Васильев Ю. М., Гельфанд И. М. и др., «Природа», №6, 13 и №7, 24, 1969.
6. Бернштейн Н. А., Проблемы кибернетики, вып. 6, 101, 1961.
7. Бауэр Э. С., Теоретическая биология, ВИЭМ, М.– Л., 1935.
8. Коган А. Б., ДАН СССР, 154,1231, 1964.
9. Режабек Б. Г., Материалы III Всесоюзной научной конференции по нейрокибернетике, Ростов-на-Дону, 1967.
10. Блехман И. И., Синхронизация динамических систем, М.,«Наука»1971.
11. Экклс Дж.,Физиология нервных клеток, ИЛ, М., 1969.
12.Ильинский О. Б., «Механорецепторы. Вопросы физиологии сенсорных систем», «Наука», Л., 1967.
13. Нуden H., Proc. Fourth Inter. Congr. Biochemistry, 3, 64, L., Pergamon Press, 1959.
Поступила в редакцию 8.VI.1972
О НЕКОТОРЫХ ОСОБЕННОСТЯХ СТАТИЧЕСКИХ И ДИНАМИЧЕСКИХ ХАРАКТЕРИСТИК НЕЙРОРЕЦЕПТОРНОЙ КЛЕТКИ
МАТЕРИАЛЫ XVII НАУЧНОЙ КОНФЕРЕНЦИИ ФИЗИОЛОГОВ ЮГА РСФСР
г. Ставрополь, 1969
I ТОМ, с.22—23
В литературе сложился взгляд на нейрон, как на аналоговоимпульсный преобразователь информации. Во многих работах авторы считают, что частота импульсной серии полностью определяется значением генераторного потенциала /ГП/. Эта же точка зрения принята в ряде математических моделей /Попов, 1965, 1967/.Однако реальный процесс преобразования информации следует разбить на два этапа: I) возникновение ГП в ответ на внешнее воздействие и 2) возникновение серии импульсов в аксонном холмике, зависящее от соотношения между ГП и критическим уровнем деполяризации / КУД/ Последний, в свою очередь) не есть величина постоянная, а может зависеть как от величины ГП, так и от скорости его изменения.
В качестве объекта экспериментальных исследований был выбран сенсорный нейрон рецептора растяжения речного рака /СНРР/. Были изучены статические характеристики, отражающие зависимость частоты импульсной активности от степени растяжения. Характеристики линейны в начальной части, затем в диапазоне 30—40 имп/сек наступает насыщение, после чего следует срыв.
Известно, что частота срыва не является предельной для нервного волокна рецептора, т.к. деполяризуя клетку можно достичь частоты 120 и более имп/сек. Поэтому можно считать, что при медленном растяжении частота разряда отражает изменения уровня ГП, который, в свою очередь, связан с избирательным повышением проницаемости сенсорной мембраны к ионам натрия в силу изменения состояния структурированной воды в порах участков мембраны, подвергаемой растяжению. /Режабек, 1966/. Однако при переходе к изучению динамических характеристик ситуация существенно меняется. При исследовании по методу Фурье вместо ожидаемых плавных изменений частоты с некоторым сдвигом по фазе, типичных для линейных систем, обнаружены периодические «вспышки» импульсов. Этот факт может быть сопоставлен с «биокибернетическим законом однонаправленной чувствительности» Клайнса и указывает на существенную нелинейность объекта по отношению к производной сигнала.
Изучение зависимости формы характеристик от скорости растяжения показывает закономерный сдвиг кривых при нарастании скорости воздействия. При быстром сжатии импульсы исчезают уже в том диапазоне, в котором при медленном сжатии нейрон работает с частотой 20—25 имп/сек. Изменение частоты после резкого растяжения, представленное в логарифмическом масштабе времени, четко выражено двумя отрезками с различной степенью наклона. Это говорит о двух процессах, ведущих к снижению частоты. Один из них, вероятно, связан с вязко-эластическими процессами в мышце, а второй следует связать с изменениями КУД генеративной мембраны /аккомодацией/. Представления об изменении пороговых свойств зависимости от скорости изменения внешнего воздействия нам представляется интересным связать с общебиологической концепцией «устойчивого неравновесия» живых систем, выдвинутой в работах Э. Бауэра /Бауэр, 1935/.
ДЕЙСТВИЕ ОБЛУЧЕНИЯ РАЗЛИЧНЫМИ УЧАСТКАМИ СПЕКТРА НА ИМПУЛЬСНУЮ АКТИВНОСТЬ РЕЦЕПТОРНОГО НЕЙРОНА РЕЧНОГО РАКА
МАТЕРИАЛЫ XVII НАУЧНОЙ КОНФЕРЕНЦИИ ФИЗИОЛОГОВ ЮГА РСФСР
г. Ставрополь, 1969
I ТОМ, с. 31 – 32. В. В. Радченко, Б. Г. Режабек
Нами собрана установка, позволяющая проводить микрооблучение клетки, допускающая формирование светового зонда до 3 микрон в диаметре (Режабек,1966). Опыты проводились на изолированном рецепторе растяжения речного рака.
Интактный рецептор способен длительно (до нескольких часов) поддерживать постоянный уровень ритмической активности с частотой 2—0 имп/сек.
Облучение участком спектра 2600—2800Ǻ (комбинация хлорбромного фильтра с фильтром УФС-I) приводит к замедлению ритмики и характерным перебоям в импульсной активности. Эффект обратим: на одной клетке опыт можно повторить 6—10 раз (при длительности облучения 1 мин. и перерывах между облучениями до 10 минут).
Иное действие оказывают лучи, выделенные светофильтрами с полосой пропускания вблизи 3660Ǻ и 3130Ǻ. Обе эти области оказывают возбуждающее действие на нейрон. Импульсная активность через 10 сек после облучения увеличивается на 30—40% и удерживается на новом уровне.
До сих пор речь шла о действии УФ спектра. Существенно необратимое действие оказывает полный спектр. Частота импульсов за 10 сек доходит до 80—100 имп/сек, после чего наступает срыв активности, очевидно, связанный с гибелью клетки. —
Клетку можно сенсибилизировать к облучению видимым светом путем прокрашивания ее различными люминофорами, имеющими сродство к определенным молекулярным компонентам цитоплазмы.
Одним из наиболее изученных люминофоров является акридиновый оранжевый (АО), применяющийся в цитохимии для выявления нуклеиновых кислот. На рецепторе, прокрашенном АО (аналогичный аффект наблюдался с уранином) наблюдался интересный эффект резкого торможения (через 1—2 сек после облучения). Эффект полностью обратим, причем восстановление исходного уровня активности также происходит очень быстро (за 2—3 сек). Нужно заметить, однако, что этот эффект связан скорее всего с действием не световых, а тепловых лучей, т.к. освещение с тепловым фильтром не вызывает подобного эффекта.
ИДЕИ Э.С.БАУЭРА И ПРОБЛЕМЫ БИОФИЗИКИ
МОСКОВСКОЕ ОБЩЕСТВО ИСПЫТАТЕЛЕЙ ПРИРОДЫ ДОКЛАДЫ МОИП «ОБЩАЯ БИОЛОГИЯ»
Доклад прочитан 20 мая 2006 года
Творчество замечательного советского биолога Э.С.Бауэра привлекает в начале третьего тысячелетия все большее внимание биофизиков, биологов и философов. Эрвин Симонович Бауэр является одним из тех ученых, которые не боялись заглядывать далеко вперед и, опираясь на логику и мощную интуицию, создавать пути для решения фундаментальных проблем биологии, намного опережая уровень знаний своего времени.
Сегодня его книга переиздана уже несколько раз, и, что особенно интересно, даже в издательстве «R&C dynamics», специализированном на издании классиков науки в области физики и математики. Отнюдь не случайно великий русский физиолог А.А.Ухтомский сделан на своей книге, подаренной Бауэру, надпись: «Эйнштейну в биологии»!
Бауэр, как и Эйнштейн, дает нам пример того, что такое гениальность в науке, подтверждая точную мысль А.С.Пушкина «Гений – парадоксов друг»
Ведь гений, как известно, смотрит на вещи, которые видели все, но видит то, чего до него не видел никто другой. Действительно, главная идея Бауэра об «устойчивом неравновесии»живой материи ничуть не менее парадоксальна, чем мысль о том, что скорость света не зависит от системы координат.
До Бауэра все физиологи были уверены в том, что хрестоматийные опыты Рубнера с измерением теплоты, выделяемой в животным в калориметре при «сжигании» пищи, являются подтверждением того, что и для живых существ закон сохранения энергии выполняется так же, как и для неживых физических систем. Бауэр же увидел в этом эксперименте, принципиально важном для борьбы с витализмом, но тривиальном с точки зрения термодинамики, нечто совершенно иное – парадокс свободной энергии живого вещества.
Фундаментальным основанием термодинамики является Первое Начало, согласно которому внутренняя энергия любого физического тела (системы) может быть изменена только двумя способами: с помощью работы δА или с помощью передачи тепла δQ.
Математически Первое Начало термодинамики выражается уравнением
ΔЕ = – δА + δQ
Знак «-» перед приращением работы связан с тем, что работу считают положительной, если тело (система) совершает её над окружающими телами, и отрицательной, если работа совершается над телом. Приращение внутренней энергии ΔЕ является полным дифференциалом, а внутренняя энергия Е – функцией состояния, но ни δА, ни δQ полными дифференциалами не являются, что подчеркивается использованием греческой, а не латинской буквы для обозначения их приращений. Но опыты Рубнера свидетельствуют о том, что ΔЕ = δQ, и, следовательно, δА равно нулю!