




Полная версия
Изыскания о древней истории Руси-России
Рассмотренное выше положение континентов подтверждается данными фаунистического районирования, согласно которым суша Земли разделяется на четыре фаунистических царства: Арктогею, Палеогею, Неогею, Нотогею. Антарктическая суша, населённая преимущественно морскими животными не входит ни в одно из царств. Арктогея («северная земля») с центром группирования на Русской платформе включает также Голарктическую, Индо-Малайскую, Эфиопскую области и занимает Евразию (без Индостана и Индокитая), Северную Америку, Северную Африку (включая Сахару). Животный мир Акртогеи характеризуется общностью происхождения. В Арктогее обитали только плацентарные млекопитающие.
Неогея («новая земля», более поздняя по времени, образовавшаяся из продуктов распада Гондваны) занимает Южную, Центральную Америку от Нижней Калифорнии и южной части Мексиканского нагорья на севере до 40° ю.ш. на юге и прилежащие к Центральной Америке острова. Распространены плацентарные. Нотогея («южная земля») занимает Австралию, Новую Зеландию и острова Океании. Длительная изоляция Нотогеи привела к формированию фауны, богатой эндемиками (изолированные виды). Число плацентарных млекопитающих относительно невелико: мышиные, рукокрылые, псовые. Палеогея занимает главным образом тропические районы Восточного полушария. Для Палеогеи характерны группы животных древней фауны Гондваны – ее Бразильско-Африканского континента: страусы, двоякодышащие рыбы, черепахи, а также хоботные, человекообразные обезьяны, хищные и др.
Указанное выше распространение фауны заостряет наше внимание на особой концентрации плацентарных млекопитающих – в пределах Арктогеи с центром на Русской равнине. Первые плацентарные известны из раннего мела (мел – 135 – 65 млн. лет назад), отложения которого занимают обширные площади на Русской платформе. Между тем, плацентарные млекопитающие, являются, во-первых, живородящими, а во-вторых, характеризуются наиболее высокой организацией и эколого-морфологическим разнообразием – головной мозг имеет сильно развитые большие полушария, которые соединены мозолистым телом; эмбриональное развитие протекает с образованием плаценты. К плацентарным принадлежит и человек. Преобладание плацентарных на территории Арктогеи даёт нам надёжные основания для утверждения именно этой области в качестве наиболее вероятной прародины человека.
Природные условия той или иной части Земли способствуют возникновению жизни – как животной, так и растительной. В пищевой цепи не могут существовать исключительно плотоядные или исключительно травоядные. А, следовательно, для того чтобы уверенно указать на присутствие жизни в том или ином регионе, следуют установить хотя бы одну из этих составляющих – либо останки животных, либо останки растений. Последние, разлагаясь, образуют угли. Отсюда следует, что, определив размещение месторождений углей на планете, мы одновременно определим и места проистечения жизни. Сделаем экскурс в древнюю историю Земли в том её аспекте, который касается условий образования каменного угля.
Угли образуются из остатков высших наземных растений. Отложение их происходило преимущественно в болотах, занимавших низменные побережья морей, заливов, лагун, пресноводных бассейнов (озёр и рек). Накапливавшийся растительный материал в результате биохимического разложения перерабатывался в торф; при этом значительное влияние оказывали обводнённость и химический состав водной среды. Торфообразование и торфонакопление завершались перекрытием торфяника осадками, образующими породы кровли. Диагенетические и биохимические процессы приводили к превращению торфа в бурый уголь. В результате длительного воздействия повышенных температур и давления бурые угли преобразовывались в каменные угли, а последние – в антрациты.
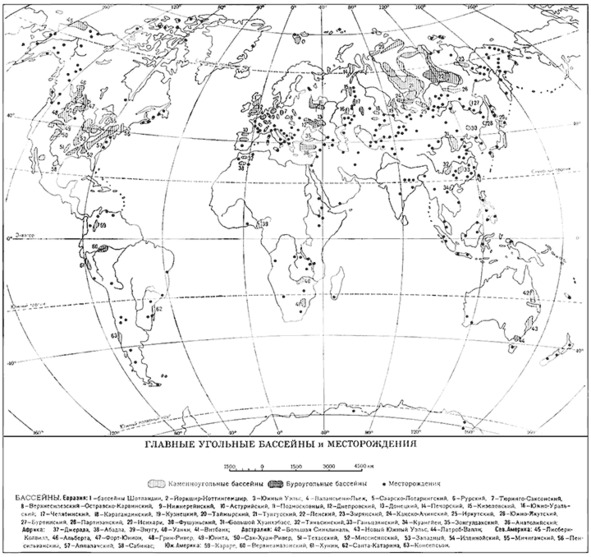
Рис. 1
На карте «Главные угольные бассейны и месторождения» (Рис. 1) отчётливо определяются основные зоны образования углей. Они сосредоточены почти исключительно в Северном полушарии и расположены между 40° и 50°. На Евразийском континенте месторождения углей имеют сплошную протяжённость от Западной Европы через Русскую равнину и до Дальнего Востока и долины реки Хуанхэ. На Русской равнине имеется ряд угольных бассейнов, например, Подмосковный угольный бассейн. Напротив, ни в Египте, ни в Месопотамии, ни в Индии месторождений углей практически нет. Анализируя такое расположение углей, приходим к следующим выводам:
Начавшееся с палеозоя активное заселение растениями территорий Земли происходило неравномерно;
Преимущественные зоны произрастания были сосредоточены в Северном полушарии на территории древнего материка Лавразия, состоящего из древних платформ – Русской, Сибирской, Китайской и Северо-Американской;
На территории другого древнего континента – Гондваны, состоящего из острова Мадагаскар, полуострова Индостан (южнее Гималаев), Африканской (без гор Атласа), Южно-Американской (к востоку от Анд), Антарктической платформ, а также Аравии и Австралии, уровень растительности либо отсутствовал полностью, либо был крайне незначительным.
«Таким образом, очевидно, что Северное полушарие в виде древнего материка Лавразии по сравнению с южной Гондваной ещё в палеозое имело более привлекательные условия для формирования жизни. Что нашло своё отражение в возникновении и распространении основных масс высших растений, дошедших до нас в виде залежей углей. Эта картина полностью повторяется и в настоящее время в виде полной аналогии распространения плодородных почв, основанных на тех же гуминовых соединениях».
Формирование материков, климата и современной флоры и фауны.
Отмечаем, что на всех материках наблюдался активный вулканизм, своими выбросами и другими последствиями негативно отразившийся на жизнедеятельности местных форм флоры и фауны. Исключением из этого шлейфа активности являются только часть Северной Америки, Антарктида и Русская платформа, сформированная на стабильной архейской (3,5 млрд. л.н.) платформе. Ввиду этого возможное перемещение фауны, включающей предков обезьян и далее человека, с материка на материк до 68 млн. л.н. не происходило. Только в конце этого периода к югу Русской платформы присоединились Азия и осколки праафриканской Гондваны – Индия и Аравия, не имеющие изначально обезьян.
Далее в истории Земли наступил палеогеновый период (с 68 млн. л.н.), в котором происходили крупные тектонические движения. В западной части Северной и Южной Америки продолжался рост горных сооружений Кордильер и Анд, сопровождаемый активной вулканической деятельностью. Во второй половине палеогенового периода (с 45 млн. л.н.) в разных частях планеты происходили складчатость и горообразование, что привело к возникновению двух цепей гористых островов, огибающих с юга и севера Средиземное море – сформировались Европа и Северная Африка с продолжающимся вулканизмом (на Балканском полуострове, в Закавказье, в Малой Азии).
Палеогеновая трансгрессия достигла максимума к концу эоцена (37,2 млн. л.н.), к этому времени море затопило южную половину Восточно-Европейской платформы, Туранскую и Западно-Сибирскую плиты, Закавказье, Среднюю и Южную Европу, север Африки и т. д. Между Русской платформой и Африкой и в южной половине Азии существовала крупная геосинклинальная система, которая простиралась от современных Пиренеев до Бирмы. Сообщению животного и растительного миров Африки и Русской платформы препятствовала полоса воды, шириной около 2500 км, а Русской платформы и Азии – более 1000 км. Причём, Африка и Азия являлись зоной повышенного вулканизма, а Русская платформа и Северная Америка – зоной тектонической стабильности. Европа и Северная Африка – не существовали.
В начале олигоцена (37,2 млн. л.н.) произошли тектонические движения, приведшие к регрессии моря (за исключением Средней Европы, Карибского моря, Мексиканского залива и др.). К началу палеогенового периода вымерли динозавры, и широкое распространение получили млекопитающие, в палеоцене представленные примитивными формами, а с эоцена (конец 37,2 млн. л.н.) появляются насекомоядные, грызуны, приматы, хищные, копытные, хоботные, китообразные и сиреновые.
Плацентарные делятся на четыре надотряда: xenarthra, лавразиотерии, афротерии, euarchontoglires. Xenarthra содержит отряд неполнозубых, происхождение – вероятно, Южная Америка. К афротериям относят немногочисленную группу животных – трубкозубые, даманы, сирены, хоботные, прыгунчиковые и тенрекообразные, прародина – вероятно, Африка. Интересующие нас обезьяны, как и сам человек – входят в сестринскую группу euarchontoglires надотряда лавразиотериев. Последний общий предок обеих групп жил от 85 до 95 миллионов лет назад на континенте Лавразия (сложился 500 – 440 млн. л.н. из древних платформ – Русской, Сибирской, Китайской и Северо-Американской, см. выше). В надотряд euarchontoglires входят грызунообразные, включающие грызунов и зайцеобразных, а также euarchonta, к которым относятся тупайеобразные, шерстокрылы и приматы. В надотряд лавразиотериев входят насекомоядные, рукокрылые, ящеры, хищные, непарнокопытные, парнокопытные и китообразные.
Как показатели достижений эволюции выходцев с территории Русской платформы (лавразиотериев) можно привести рукокрылых – единственная группа млекопитающих, освоившая воздушную стихию, и синего кита – являющего не только самым крупным млекопитающим, но и самым крупным известным животным. Немудрено, что именно выходцы лавразиотериев дали миру человека.
Большинство родов и многие виды растений неогена (23,0 – 1,81 млн. л.н.) (особенно плиоцена, 5,33 – 1,81 млн. л.н.) существуют доныне, хотя географическое распределение их во многих случаях изменилось. В начале неогена (с 23,0 млн. л.н.) климат Северного полушария был более тёплым и влажным, чем в современную эпоху. В Сибири преобладали широколиственные леса, в Западной Европе в областях, расположенных на сравнительно высоких широтах, росли пальмы, лавры и мирты. К концу неогена (1,81 млн. л.н.) Сибирь была покрыта хвойной тайгой, хотя по долинам рек ещё произрастали грецкие орехи. В Западной Европе к концу неогена вечнозелёные формы были оттеснены к побережьям Средиземного моря, сменившись севернее листопадными и хвойными лесами. Процессы постепенного похолодания и увеличения сухости климата устанавливаются по ископаемым флорам Северного и Южного полушарий. Ископаемая флора тропического пояса очень мало отличалась от современной. На равнинах Средней Азии и Казахстана близкая к современной степная и пустынная растительность также существовала с начала неогенового периода.
В Альпийской складчатой области Южной Европы и Юго-Западной Азии в конце палеогена началось поднятие многочисленных горных хребтов (Альп, Карпат, Балкан, Динарских гор, Апеннин, Кавказа, Крыма, гор Понта и Тавра, Загроса, Белуджистана, Гималаев и др.). Рост гор сопровождался образованием межгорных и краевых впадин. Вдоль разломов магма проникала на поверхность, изливаясь в виде лавовых покровов и образуя вулканические конусы. Главными центрами вулканизма были Апеннинский полуостров, Малая Азия, юг Балканского полуострова, Кавказ. В конце неогенового периода произошло образование глубоких впадин внутренних морей – Лигурийского, Тирренского, Ионического, Чёрного, Каспийского, а также Адриатического, Мраморного.
Начало неогена на территории Европы сопровождалось резким обновлением наземной фауны: вымерли сумчатые, древние хищники – креодонты, многие группы примитивных копытных. Их сменили представители многих новых семейств, в большинстве своём существующих доныне: древние виды медведей, барсуков, гиен, первые хоботные, предки лошадей – анхитерии, первые свиньи, антилопы, олени, быки, овцы, человекообразные обезьяны. Среди них появились, в частности, обезьяны. Млекопитающие Северной Америки в начале неогена развивались обособленно и были значительно менее разнообразны. В середине неогена (верхний миоцен, ок. 5,5 млн. л.н.) между континентами Европы, Азии и Северной Америки установилась связь по суше (вероятно, в области Берингова пролива), что привело к большим миграциям млекопитающих и дальнейшему их развитию. В это время на обширных пространствах Европы и Азии распространяется очень однородная фауна степного типа, наиболее характерным и типичным представителем которой была трёхпалая лошадь – гиппарион. В несколько более молодых отложениях неогена встречаются наиболее древние остатки ласки, росомахи, настоящих лошадей и слонов. Конец неогена характеризуется в Европе исчезновением многих родов животных, которые, однако, продолжают существовать в современных странах тропического пояса. В миоцене и плиоцене обезьяны были широко распространены в Европе, Азии, Африке. Для антропогенеза интерес представляют миоценовые (23,0 – 5,33 – млн. л.н.) дриопитеки как возможные общие предки человека и современных человекообразных обезьян. Расцвет высших антропоморфных обезьян относится к плиоцену и началу плейстоцена (5,33 – 1,8 млн. л.н.).
Из всего вышеотмеченного можно сделать следующие выводы:
Русская равнина (без современной Европы) сформирована на древнейшей из возможных платформ – архейской (3,5 млрд. лет), что является залогом тектонической стабильности региона;
Изначально Африка и Русская платформа не вступали в геологический контакт, будучи разделёнными тысячами километров геосинклинальных вод;
Вследствие многомиллионолетней стабильности Русской платформы, плацентарные, включая приматов, образовались с 23 млн. л.н. и развились на территории Русской платформы, являющейся в указанный период центром формирования материка Лавразия. Африка в указанный период тектонировала, была бедна видами и не имела приматов;
Активные тектонические процессы происходили и на территориях, обычно указываемых местом проживания человекообразных обезьян – Северная Африка и районы Средиземноморья, – не прекращаясь, вплоть до конца миоцена (5,33 млн. л.н.). Только к этому времени сформировалась Европа и западная часть Азии, просторы которых стали пригодны для постоянного проживания животных;
Начиная с неогена, в Средней Азии и Казахстане установились современные природные условия, не способствующие развитию жизни;
До последнего времени горные системы Северной Африки, Европы и центральной Азии не прекращали своего роста, чем делали невозможным проживание и, тем более, развитие местного цивилизованного существа, такого, как, например, человек;
Разнообразие русской (европейской) фауны является результатом долговременных стабильных природных условий, свободных от тектонической активности. В то же время это разнообразие видов является лучшим показателем эволюционного развития животного мира (включая человека), осуществлённого на достаточно небольшом участке суши (Русской равнине) не путём межконтинентальных миграций, а в силу длительности существования и постепенной дифференциации.
Привязка возможных мест появления человека к палеоматерикам.
Ранее мы выявили интересную особенность современного расположения рас – центры концентрации каждой из них, во-первых, не совпадают, во-вторых, равноудалены друг от друга (центры даны приблизительно и с ориентировкой на современное размещение рас). Теперь мы можем добавить и третью особенность – ареал каждой из рас совпадает с соответствующим фаунистическим районом. Европеоидная раса – с Арктогеей. Монголоидная раса – с Неогеей. Австралоидная раса с Нотогеей. Негроидная раса – с Палеогеей.
Согласно современным данным, многие учёные объединяют негроидную и австралоидную расы в одну большую негро-австралоидную расу, родиной которой, согласно геологическим данным, является палеоматерик Гондвана. Эти же данные указывают в качестве прародины европеоидной расы Арктогею, которая соответствует Русскому палеоконтиненту. Очевидно, процесс смешивания рас говорит нам о более поздних временах в истории цивилизации – когда первоначально «чистые» представители одних, ранее изолированных рас в процессе своего расширения начали осуществлять регулярные и продолжительные контакты с представителями других, ранее изолированных рас. Следует отметить: если такой процесс смешения рас продолжается и в настоящее время, значит, он ещё не закончен. Вывод – первоначально существовало разделение рас (см. выше).
В более ранние исторические времена существование «чистых» рас ограничивалось такими областями, которые, как мы показали выше, были равноудалены от подобных областей других рас и соответствуют областям древних континентов. Очевидно, именно в силу географической разобщённости в период позднего палеолита завершился начавшийся еще на первых ступенях человеческой истории процесс расообразования, и оформились три основные человеческие расы – европеоидная, негроидная и монголоидная. Скорее всего, образование рас закончилось к началу позднего палеолита – 35 тыс. до н.э., так как к его концу (11 тыс. до н.э.) уже полным ходом шёл процесс перемешивания рас, и, как следствие, началось возникновение народов и языков.
Место возникновения негроидной расы найти не представляет труда – «древнейшее население Сахары было негроидным», то есть – это Палеогея, африканская составная часть Гондваны. Позднее, по данным Дьяконова И. М. и Милитарёва А. Ю., практически светлокожие европеоиды атлантического типа засвидетельствованы в египетских росписях конца 2 – начала 1 тыс. до н.э. у западных границ Египта. «Здесь вряд ли следует искать родину какой бы то ни было значительной популяции. Кроме того, имея в виду, европейское происхождение светлокожих европеоидов, правильнее предположить, что в Восточную Сахару они попали уже вторично». Эти данные подтверждает и другой источник: «…уже с появлением скотоводства в Сахару проникли народы средиземноморской и эфиопидной рас. В 4-м тыс. до н.э. в долине р. Нил постепенно складывается древнеегипетское государство. Страны к западу от Египта населяли ливийские племена – светловолосые, называемые египтянами темеху (тиумах), и темноволосые и смуглые – техену». Следовательно, Африка не является местом рождения европеоидов, а «приняла» их только значительно позже 11-го – 10-го тыс. до н.э.
Место возникновения монголоидной расы определить также не представляет труда – американское «коренное население – индейцы – принадлежат к монголоидной расе», «обнаруживая особое сходство с древним протомонголоидным типом», то есть – это Неогея, Лавренция, американская составная часть Лавразии. Однако монголоидная раса распространена и в Азии. Она делится на несколько малых рас: североазиатскую (8 – 3 тыс. до н.э.), арктическую (туземных обитателей Гренландии и вообще северных полярных стран, до 78° с.ш., особенно у берегов Америки), южно-азиатскую (юго-восток Азии), дальневосточную, американскую (протомонголоиды), центрально-азиатскую (с 3 – 4 веков, путём смешивания с европеоидами).
Собственно монголоидные расы в Азии подразделяются на две главные группы – континентальную и тихоокеанскую. В составе континентальных монголоидов выделяются североазиатская и центрально-азиатская расы. Промежуточное положение между континентальными и тихоокеанскими монголоидами занимает арктическая (эскимосская) раса. Северные группы тихоокеанских монголоидов объединяются в дальневосточную, или восточноазиатскую, расу. Наиболее «чистую» монголоидную расу мы обнаруживаем на Американском континенте и вблизи Берингова пролива, разъединяющего Азию и Аляску. К югу и западу от этих азиатских областей присутствуют только смешанные расы.
«Таким образом, можем сделать вывод, что негроидная раса сформировалась на юге Африки, а монголоидная – на территории Америки. Их возникновению в оригинальном виде способствовали местные особенности условий жизни. Достигнутые ими уровни цивилизации были обусловлены также местными условиями жизни. А их более раннему проникновению на территорию Евразии и далее, на территорию Русской равнины, препятствовали естественные морские преграды, делавшие древние континенты – Арктогею, Неогею, Нотогею, Палеогею – территориально разобщёнными».
Как не казалось бы это невероятным, определить место возникновения европеоидной расы также не представляет труда – «почти всё население Европы относится к большой европеоидной расе, которая делится на несколько малых рас». Отметим, что ни в Африке, ни в Австралии, ни в Америках, ни в монголоидной части Азии не отмечено присутствие человека европеоидной расы временем, ранее 10-го тыс. до н. э. В Африке и Америке, как мы показали выше, имелись центры возникновения собственных рас.
Между тем, Европа – это западная часть материка Евразии, простирающаяся на востоке до гор Урала. Но основой Европы (больше половины её территории) является Русская платформа – в древности Русский континент (см. выше). Русская платформа заселена с 70 – 50-го тыс. до н.э. (стоянки Сунгирь, Маркина гора и др.) практически исключительно русским (называем по имени Русской равнины) народом, из состава которого украинцы и белорусы окончательно выделились в самостоятельные объединения только в 20-м века н. э. Та часть Европы, которая расположена западнее Русской платформы, населена, начиная с 10 – 5 тыс. до н.э., а особенно активно – с начала «нашей» эры, расселяющимися русскими племенами, говорящими на т.н. индоевропейских языках.
Итак, европеоидная раса подразделена на три главные группы: южную – со смуглой кожей, тёмными глазами и волосами; северную – со светлой кожей, значительной долей серых и голубых глаз, русых и белокурых волос; промежуточную, для которой характерна среднеинтенсивная пигментация. «У русских – черты северной ветви европеоидной расы». Антропологи всех светлых европеоидов подразделяют на северо-западных (Атланто-балтийская раса) и северо-восточных (Беломорско-балтийская раса).
С точки же зрения академической науки, европеоидные расы сформировались первоначально в Юго-Западной Азии, Северной Африке и Европе. Однако в первых двух регионах варианты возраста рас 10 – 1 тыс. лет, смешанные с негроидами. В Европе (на Русской платформе) – образовалась изначальная русская раса (то, что сейчас называется европеоидной большой расой) возрастом более 50-и тыс. лет (по датировке европеоидов с русских стоянок, что соответствует и данным ДНК-генеалогии). На территории Скандинавских стран, Великобритании, Ирландии, Исландии, Нидерландов, на севре Германии, а также в Эстонии и Латвии распространена атланто-балтийская раса (собственно – русская раса). Отметим, например, что заселение Англии состоялось кельтами с 8 – 7 веков до н.э., которые первоначально в 1-й половине 1-го тысячелетия до н.э. обитали в бассейнах Рейна, Сены и Луары и верховьях Дуная – то есть на Русской равнине. Население центральных областей Западной Европы и Европейской части России (собственно Руси) относится главным образом к различным вариантам среднеевропейской (русской) расы.
Отметим: на протяжении пути – Урал-Европа – лежала огромная территория Центральной части Руси – Русская равнина. Межрасовые контакты зауральских монголоидов и европейцев не только не могли миновать население Русской равнины, которое было многочисленно и полностью составляло саму европеоидную расу, но и состоялось непосредственно на территории Руси. Следует особо отметить, что находки древнего человека, сделанные на территории Русской равнины (реконструированные образы), как нельзя лучше иллюстрируют это.
Так, заключение М.М.Герасимова «по сунгирьскому человеку – Homo sapiens, европеоид, 55 – 57 лет. Рост 176 – 177 см. Физически сильный, мускулистый». А девочка 10 – 11 лет, найденная на той же стоянке, обладает вполне современным видом. И, по результатам специально проведённого группой экспертов опроса, её сочли современной и даже весьма привлекательной почти сто процентов опрошенных. Более того, они безапелляционно идентифицировали её как «обычную современную русскую девушку». То же касается и вида костёнковца (стоянка Костёнки на Дону, 25 тыс. лет назад) – «вполне современный мужчина лет сорока – сорока пяти из обычной русской деревни». Очевидно, что за 30 тысяч лет, разделяющих сунгирийцев и костёнковцев от современных русских, физический тип человека русской (европеоидной) расы не претерпел значительных изменений. В силу чего возникает вопрос: если за 30 тыс. лет этого не произошло, то с какой стати физический тип человека должен были измениться за сопоставимые периоды – 50, 100, 200 тысяч лет – от потомка обезьяны – при полной изоляции региона от остального мира.
«Таким образом, можем сделать вывод, что европеоидная раса изначально сформировалась на территории Руси – на территории Арктогеи, то есть древнего Русского континента, занимающего всю площадь древнейшей в мире Русской платформы и отделённого от негроидов и монголоидов естественными водными и горными преградами».
При этом возникает естественный вопрос, а какое отношение ко всему этому может иметь Гиперборея, которая также позиционируется на Русской равнине, на ее северо-западном побережье. Утверждается, что Гиперборея была населена выдающимися белыми людьми, располагалась севернее по отношению к Греции. Геологам хорошо известен Русский материк, носивший название Арктогея (Аркт (о) + Гея – медвежья земля). Русский материк существовал в мезозое – палеозое. Его очертания соответствовали современной Русской платформе в границах – дельта Дуная, Днестр, Висла, Норвежское море, Баренцево море, реки Печора, Уфа, Белая, север Каспийского моря, дельта Волги, север Чёрного моря. Центр платформы – город Владимир в междуречье Оки и Волги. В начале палеозоя (ордовикский период, 500 – 440 млн. лет назад) Русский материк (Арктогея) стал центром формирования в Северном полушарии континента – Лавразия (в него вошли платформы: Русская, Сибирская, Китайская и Северо-Американская). Отметим, что Арктогея с центром группирования на Русской платформе включала также Голарктическую, Индо-Малайскую, Эфиопскую области. Животный мир Арктогеи характеризовался общностью происхождения. В Арктогее обитали только плацентарные млекопитающие, из которых и могли образоваться люди. Напротив, в Гондване, широко рекламируемой в качестве родины человечества, плацентарных не было, поэтому теория «африканского происхождения человека» в этой части не корректна.







