


Полная версия
Тест на ДНК. С чего все начиналось? О наследственности, изменчивости и эволюции



Если обозначить через А один из пары константных признаков, например доминирующий, через а – рецессивный и Аа – гибридную форму, в которой оба признака соединяются, то получается выражение:
А + 2Аа + а,
обозначающее ряд развития (Entwicklungsreihe) потомков гибридов для каждой пары различающихся признаков.
Сделанное Гэртнером, Кёльрейтером и другими открытие, что гибриды обладают склонностью возвращаться к родоначальным формам, подтверждается вышеупомянутыми опытами. Можно показать, что число гибридов, явившихся в результате одного оплодотворения, начинает от поколения к поколению значительно отставать от числа сделавшихся константными форм и их потомков, но совсем исчезнуть гибриды не могут. Если мы допустим в среднем для всех растений во всех поколениях одинаковую плодовитость и примем дальше, что каждый гибрид образует семена, из которых наполовину опять выходят гибриды, наполовину же константные формы с поделившимися поровну обоими признаками, то численные отношения для потомков в каждом поколении вытекают из нижеследующей сводки, где А и а опять образуют оба исходных признака и Аа – гибридную форму. Для краткости можно предположить, что каждое растение образует в каждом поколении только 4 семени.
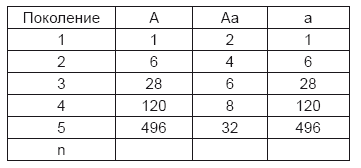
Выровненные в пропорциях
A: Аа: а
1: 2: 1
3: 2: 3
7: 2: 7
15: 2: 15
31: 2: 31
2n–1: 2: 2n–1
В 10-м поколении, например, 2n – 1 = 1023. Следовательно, на каждые 2048 растений, которые выходят из этого поколения, приходится 1023 с константно доминирующим признаком, 1023 с рецессивным и только 2 гибрида.
Потомки гибридов, в которых соединено несколько различающихся признаков
При обсуждавшихся выше опытах употреблялись растения, которые отличались только в одном существенном признаке. Ближайшая задача состояла в том, чтобы исследовать, действителен ли найденный закон развития каждой пары различающихся признаков также и в том случае, когда несколько различных особенностей соединяются в гибридах при оплодотворении.
Что касается общего вида гибридов в этом случае, то опыты согласно показывают, что они ближе к тому из двух основных растений, которое имеет большее число доминирующих признаков. Если, например, семенное растение имеет короткую ось, верхушечные белые цветы и простые выпуклые бобы, пыльцевое растение, наоборот, – длинную ось, пазушные лилово-красные цветы и бобы с перехватами, то гибрид только формой боба напоминает семенное растение, в остальных же признаках он согласуется с пыльцевым растением. Если один из двух основных видов имеет только доминирующие признаки, то гибрид совершенно или почти не отличим от него.
Были проведены два опыта с большим числом растений. В первом опыте исходные растения отличались формой семян, окраской белка и цветом семенной кожуры. Опыты с признаками семян ведут к цели проще и вернее.
С целью более легкого рассмотрения различные признаки семенного растения будут обозначены в этих опытах через А, В, С, у пыльцевого растения – а, b, с и гибридные формы этих признаков – Аа, Вb, Сс.
Первый опыт:
AB – семенное растение
A – форма круглая
B – белок желтый
ab – пыльцевое растение
a – форма угловатая
b – белок зеленый
Семена оплодотворенного [растения] были круглые и желтые, подобные таковым у семенного растения. Выращенные из них растения дали семена четырех сортов, которые часто располагались совместно в одном бобе. В общем от 15 растений получилось 556 семян, из которых было:
315 круглых и желтых,
101 угловатое и желтое,
108 круглых и зеленых,
32 угловатых и зеленых.
Все они были выращены в следующем году. Из круглых желтых семян 11 не взошли и 3 растения не дошли до образования плодов. Из остальных растений имели:
38 круглые желтые семена АВ
65 круглые желтые и зеленые семена АВb
60 круглые желтые и угловатые желтые семена АаВ
138 круглые желтые и зеленые, угловатые желтые и зеленые семена АаВb
Из угловатых желтых семян дошли до образования плодов 96 растений, из них имели:
28 только угловатые желтые семена аВ
68 угловатые желтые и зеленые семена аВb.
Из 108 круглых зеленых семян принесли плоды 102 растения, из которых имели:
35 только круглые зеленые семена Аb
67 круглые и угловатые зеленые семена Ааb.
Угловатые зеленые семена дали 30 растений с совершенно одинаковыми семенами; они остались константными аb.
Таким образом, потомство гибридов появилось в 9 различных формах и частью в очень неодинаковом числе. Если сопоставить и распределить их, то получается:
38 растений с обозначением АВ

АВ 68 растений с обозначением аВb

Все формы можно расположить в три существенно различные группы. Первая обнимает формы с обозначением АВ, Аb, аВ, аb, они имеют только константные признаки и не меняются в ближайших поколениях. Каждая из этих форм встречается в среднем 33 раза. Вторая группа содержит формы АВb, аВb, АаВ, Ааb; в одном признаке они константны, в других гибридны, и в ближайшем поколении варьируют только относительно гибридного признака. Каждая из них появляется в среднем 65 раз. Форма АаВb встречается 138 раз, гибридна в обоих признаках и ведет себя совершенно так же, как гибрид, из которого она произошла.
Если сопоставить числа, в которых появляются формы этих групп, то нельзя не признать средних отношений 1: 2: 4. Числа 33, 65, 138 дают вполне удовлетворительное приближение к отношениям 33: 66: 132.
Ряд (Entwicklungsreihe) состоит, следовательно, из 9 членов, из которых 4 представлены в нем по одному разу каждый и константны в обоих признаках; формы АВ, аb схожи с исходными видами, обе другие представляют единственные кроме них возможные константные комбинации между соединившимися признаками А, а, В, b. Четыре члена встречаются по два раза каждый и в одном признаке константны, в другом – гибридны. Один член встречается 4 раза и является гибридным в обоих признаках. Таким образом, потомки гибридов, в которых соединены двояко различающиеся признаки, получаются по формуле:
AB+Ab+aB+ab+2ABb+2aBb+2AaB+2Aab+4AaBb
Этот ряд представляет собой бесспорно комбинационный ряд, в котором связаны почленно оба ряда развития для признаков А и а, В и Ь. Полное число членов ряда получается комбинированием выражений:
Второй опыт:
ABC – семенное растение
А – форма круглая
В – белок желтый
С – кожура серо-коричневая
abc – пыльцевое растение
а – угловатая
b – белок зеленый
с – кожура белая
Этот опыт был проведен совершенно таким же образом, как и предшествующий. Из всех опытов он потребовал наибольшего количества времени и труда. Из 24 гибридов было получено в общем 687 семян, все они были с крапинами, серо-коричневые или серо-зеленые, круглые или угловатые. Из них в следующем году дошли до образования плодов 639 растений, и, как показали дальнейшие исследования, между этими растениями было:
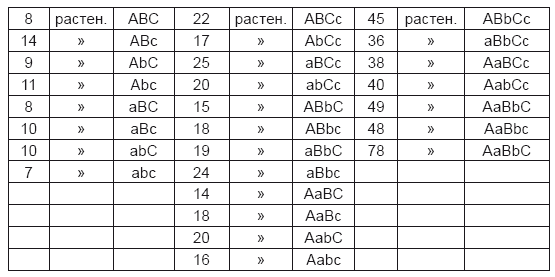
Ряд охватывает 27 членов, из них 8 константны во всех признаках, и каждый встречается в среднем 10 раз; 12 константны в двух признаках, в третьем гибридны, каждый встречается в среднем 19 раз; 6 константны в одном признаке, в двух других гибридны, каждый из них встречается в среднем 43 раза; одна форма встречается 78 раз и гибридна во всех признаках. Отношения 10: 19: 43: 78 так близко подходят к отношениям 10: 20: 40: 80 или 1: 2: 4: 8, что последние представляют несомненно истинные значения.
Развитие гибридов, если их исходные виды различны в трех признаках, следует поэтому по выражению:
АВС + АВс + АbС + Abc + аВС + аВс + abC + abc + 2АВСс + 2АbСс + 2аВСс + 2аbСс + 2АВbС + 2АВbc + 2аВbС + 2аВbс + 2АаВС + 2АаВс + 2АаbС + 2Ааbс + 4АВbСс + 4аВbСс + 4АаВСс + 4АаbСс + 4АаВbС + 8АаВbс + 8АаВbСс.
Здесь также мы имеем комбинационный ряд, в котором соединяются между собою ряды развития для признаков А и а, В и b, С и с. Выражения:
А + 2Аа + а
В + 2Вb + b
C + 2Cc + c
дают всех членов ряда. Константные соединения, которые встречаются в этих последних, соответствуют всем возможным комбинациям между признаками А, В, С, а, b, с; два из них, АВС и аbс, подобны обоим родителям.
Кроме того, было проведено много опытов с небольшим числом растений, у которых другие признаки связаны гибридно по два и по три; все дают приблизительно одинаковые результаты. Поэтому не подлежит никакому сомнению, что для всех подвергнутых опытам признаков имеет одинаковую силу следующее положение: потомки гибридов, соединяющих в себе несколько существенно различных признаков, представляют собой членов комбинационного ряда, в котором соединены ряды развития каждой пары различающихся признаков. Этим одновременно доказывается, что поведение в гибридном соединении каждой пары различающихся признаков независимо от других различий у обоих исходных растений.
Если n означает число характерных различий у обоих исходных растений, то 3n есть число членов комбинационного ряда, 4n – число индивидов, принадлежащих ряду, и 2n – число соединений, которые остаются константными. Так, например, когда исходные виды различны в 4 признаках, ряд содержит 34 = 81 член, 44 = 256 индивидов и 24 = 16 константных форм; или, что то же, между 256 потомками гибридов существует 81 различное соединение, из которых 16 константны.
Все константные соединения, которые возможны у Pisurn при комбинации семи характерных признаков, были действительно получены путем повторного скрещивания. Их число выражается в 27 = 128. Этим дается фактическое доказательство тому, что константные признаки, которые встречаются у различных форм родственной растительной группы, могут вступить путем повторного искусственного оплодотворения во все соединения, которые возможны по правилам комбинации.
Опыты, касающиеся времени цветения гибридов, еще не закончены. Насколько позволяют судить полученные доселе данные, оно стоит почти точно посередине между временем цветения семенного и пыльцевого растения, и образование гибридов относительно этого признака происходит таким же образом, как это имеет место для других признаков. Формы, которые будут выбраны для опытов этого рода, должны различаться во времени цветения по меньшей мере на 20 дней; затем необходимо, чтобы семена при посеве были посажены на одинаковую глубину в землю, чтобы достигнуть одновременного прорастания, чтобы затем во время полного цветения принимались в расчет колебания в температуре и вызванные этим частичные ускорения или замедления в расцветании. Ясно, что в этом опыте надо преодолеть различные затруднения, и он требует большого внимания.
Если мы попробуем кратко резюмировать полученные нами результаты, то мы найдем, что те различающиеся признаки, которые легко и определенно устанавливаются в опытных растениях, в гибридном соединении ведут себя совершенно одинаково. Потомки гибридов каждых двух различающихся признаков являются наполовину также гибридами, тогда как другая половина становится константной по признакам семенного и пыльцевого растения в равных частях. Если несколько различающихся признаков соединяется путем оплодотворения в одном гибриде, то потомки последнего являются членами одного комбинационного ряда, в котором соединены ряды развития для каждой пары различающихся признаков.
Полная согласованность, которую обнаруживают подвергнутые опытом признаки, допускает вполне и с полным правом предположение, что подобное же поведение присуще другим признакам, которые выступают у растений менее определенно и поэтому не могут быть применены в самостоятельном опыте. Опыт с цветочными осями различной длины дал в общем вполне удовлетворительный результат, хотя установление и расположение форм не может быть проведено с той достоверностью, которая необходима для правильных опытов.
Зачатковые клетки гибридов
Результаты, к которым привели вышеописанные опыты, побудили к дальнейшим исследованиям, которые в случае успеха должны дать объяснение свойств [женских] зачатковых (Keimzellen) и пыльцевых клеток гибридов. Важным опорным пунктом является то обстоятельство, что у Pisurn среди потомков гибридов появляются константные формы во всех комбинациях соединившихся признаков. Насколько охватывает опыт, везде подтверждается, что константные потомки образуются только тогда, когда зачатковые клетки и оплодотворяющая пыльца однородны, причем и те, и другие снабжены задатками (Anlagen), способными давать совершенно одинаковые индивиды, как это происходит и при нормальном оплодотворении чистых видов. Поэтому мы должны по необходимости принять, что при возникновении константных форм у гибридного растения объединяются совершенно одинаковые факторы. Так как одним и тем же растением и даже в одном и том же цветке этого последнего производятся различные константные формы, то будет последовательным признать, что в завязях гибридов образуется столько различных зачатковых клеток (зародышевых пузырьков) и в пыльниках столько различных пыльцевых клеток, сколько возможно константных комбинаций, и что эти зачатковые и пыльцевые клетки соответствуют по своим внутренним свойствам отдельным формам.
Действительно, теоретическим путем можно показать, что этого предположения совершенно достаточно, чтобы объяснить развитие гибридов в отдельных поколениях, если в то же время допустить, что различные виды зачатковых и пыльцевых клеток у гибридов образуются в среднем в одинаковом числе.
С целью подвергнуть эти предположения экспериментальной проверке были намечены следующие опыты: две формы, константно различные по форме семян и окраске белка, были соединены путем оплодотворения между собою.
Если обозначить различающиеся признаки опять через А, В, а, b, то были взяты:
АВ – семенное растение
А – круглая форма [семян]
В – желтый белок
аb – пыльцевое растение
а – угловатая форма [семян]
b – зеленый белок.
Семена искусственно оплодотворенных [растений] были высеяны вместе с семенами обоих исходных растений, и самые сильные экземпляры были выбраны для взаимного скрещивания. Были оплодотворены:
1) гибриды пыльцой от АВ,
2)»»» ab,
3) АВ»» гибрида,
4) ab»» гибрида
В каждом из этих четырех опытов на трех растениях были оплодотворены все цветы. Если вышеизложенное предположение было правильно, то на гибридах должны были развиваться зачатковые и пыльцевые клетки форм АВ, Аb, аВ, аb, и тогда соединялись:
1) зачатковые клетки АВ, Ab, аВ, аb с пыльцевыми клетками АВ,
2)»» АВ, Аb, аВ, ab»» ab,
3)»» АВ с пыльцевыми клетками АВ, Ab, аВ, ab,
4)»» ab»» АВ, Аb, аВ, ab.
Тогда в каждом таком опыте должны были получаться только следующие формы:
1. АВ, АВb, АаВ, АаВb,
2. АаВb, Aab, аВb, ab,
3. АВ, АВb, АаВ, АаВb,
4. АаВb, Ааb, аВb, ab.
Если даже отдельные формы зачатковых и пыльцевых клеток у гибрида образуются в среднем в одинаковом числе, то в каждом опыте приведенные четыре соединения должны появляться одинаковое число раз. Однако полного совпадения числовых отношений ожидать нельзя, так как при каждом оплодотворении, даже нормальном, некоторые клетки остаются неразвитыми или повреждаются позднее, и даже многие из хорошо развившихся семян не прорастают после посева. Другим ограничением сделанного предположения является то обстоятельство, что при образовании различных зачатковых и пыльцевых зерен имеет место лишь приближение к равенству чисел, но математической точности в пределах каждого отдельного гибрида здесь ожидать не приходится.
Первый и второй опыты были предприняты с целью испытать свойства гибридных зачатковых клеток, тогда как третий и четвертый опыты должны были решить тот же вопрос для пыльцевых клеток. Как видно из вышеприведенного сопоставления, первый и третий опыты, так же как второй и четвертый, должны дать совершенно одинаковые результаты, и этот результат должен быть частично виден уже на втором году в форме и окраске искусственно оплодотворенных семян. В первом и третьем опытах доминирующие признаки формы и окраски А и В появляются в каждом соединении, и притом частью константно, частью в гибридном сочетании с рецессивными признаками а и b, – вследствие этого они должны накладывать свой отпечаток на все семена. Поэтому, если наше предположение правильно, все семена должны были бы быть круглыми и желтыми. Во втором и четвертом опыте, наоборот, одно соединение было гибридным по форме и окраске, поэтому семена должны быть круглыми и желтыми, другое – гибридно по форме и константно в рецессивном признаке окраски, поэтому семена круглые и зеленые; третье – константно в рецессивном признаке формы и гибридно в окраске, поэтому семена угловатые и желтые; четвертое – константно в обоих рецессивных признаках, поэтому семена угловатые и зеленые. Следовательно, в обоих указанных опытах можно было ожидать четыре рода семян, а именно – круглые желтые, круглые зеленые, угловатые желтые, угловатые зеленые.
Результаты сбора полностью соответствовали поставленным требованиям.
Было получено в:
1 опыте 98 исключительно круглых желтых семян,
«94»»»»,
«31 круглое желтое, 26 круглых зеленых, 27 угловатых желтых, 26 угловатых зеленых семян,
4 опыте 24 круглых желтых, 25 круглых зеленых, 22 угловатых желтых, 27 угловатых зеленых семян.
При столь благоприятном результате нельзя было сомневаться, что ближайшее поколение принесет окончательное разрешение. Из высеянных семян в следующем году дошли до плодоношения в первом опыте 90, в третьем 87 растений; из них дали при опытах:
20.25 круглые желтые семена АВ.
23.19 круглые, желтые и зеленые семена АВb.
25.22 круглые и угловатые желтые семена АаВ.
22.21 круглые и угловатые, желтые и зеленые семена АаВb.
Во втором и четвертом опыте круглые и желтые семена дали растения с круглыми и угловатыми, желтыми и зелеными семенами АаВb.
От круглых зеленых семян были получены растения с круглыми и угловатыми зелеными семенами Ааb.
Угловатые желтые семена дали растения с угловатыми и зелеными семенами аВb.
Из угловатых зеленых семян были выращены растения, которые дали опять только угловатые зеленые семена аb.
То, что и в этих двух опытах также не проросли некоторые семена, не могло изменить чисел, добытых в предшествовавшем году, ибо каждый сорт семян дал растения, сходные между собою в отношении семян и отличные от других. Так при этом получено:
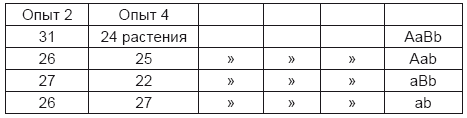
Таким образом, во всех опытах появились все требуемые при сделанном допущении формы и приблизительно в равном числе.
Для дальнейшего испытания были взяты признаки окраски цветка и длины оси, при этом выбор был сделан так, что в третьем опытном году каждый признак должен был проявиться у половины всех растений, если вышеупомянутое предположение правильно. А, В, а, b служат опять для обозначения различных признаков.

Форма Аb была оплодотворена аb, причем получился гибрид Ааb. Затем аВ было оплодотворено также ab, отсюда гибрид аВb. Во втором году для дальнейшего оплодотворения гибрид Ааb был взят как семенное растение, другой гибрид аВb – как пыльцевое растение.
Семенное растение Ааb пыльцевое растение аВb.
Возможные зачатковые клетки Аb, аb пыльцевые клетки аВ, ab.
Из оплодотворения между возможными зачатковыми и пыльцевыми клетками должны получиться четыре соединения, именно:
АаВb + аВb + Ааb + аb.
Отсюда видно, что по вышеизложенному предположению в третьем опытном году из всех растений половина должна была иметь
лилово-красные цветы (Аа)…. Члены 1-3
белые цветы (а)» 2-4
длинную ось (Вb)» 1-2
короткую ось (b)» 3-4
Из 45 оплодотворений второго года получилось 187 семян, из которых в третьем году достигли цветения 166 растений. Отдельные члены появились из них в следующем числе:
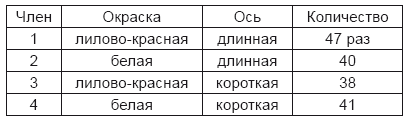
Следовательно:
лилово-красная окраска цветов (Аа) появилась у 85 растений
белая»» (а)»» 81»
длинная ось»» (Вb)»» 87»
короткая»»» (b)»» 79»
Итак, выставленное воззрение достаточно подтверждается и в этом опыте.
Для признаков формы бобов, окраски бобов и расположения цветов были также поставлены небольшие опыты, которые дали совершенно согласные результаты. Все соединения, которые были возможны при объединении различных признаков, появились в точности и приблизительно в равном числе.
Следовательно, экспериментальным путем подтвердилось предположение, что гибриды гороха образуют зачатковые и пыльцевые клетки, которые по своим свойствам и в равных количествах соответствуют всем константным формам, получающимся из комбинаций, соединенных при оплодотворении признаков.
Различие форм среди потомков гибридов, а также и численные отношения, в которых эти последние наблюдаются, находят себе достаточное объяснение в только что изложенном положении. Простейшим случаем является ряд двух различающихся признаков. Этот ряд обозначается, как уже известно, выражением А + 2Аа + а, где А и а означают формы с константно различающимися признаками и Аа – гибридную форму обоих. Он содержит в трех различных членах четыре индивида. При образовании этих последних пыльцевые и зачатковые клетки формы А и а вступают в оплодотворение в равных долях в среднем, поэтому каждая форма образуется дважды из четырех индивидов.
Следовательно, в оплодотворении участвуют:
пыльцевые клетки А + А + а + а,
зачатковые клетки А + А + а + а.
Какой из двух видов пыльцы соединится с каждой отдельной зачатковой клеткой, вполне предоставлено случаю. По теории вероятности в среднем каждая форма пыльцы А и а соединяется одинаковое число раз с каждой формой зачатковой клетки А и а; поэтому одна из двух пыльцевых клеток А встречается при оплодотворении с зачатковой клеткой А, другая – с зачатковой клеткой а; таким же образом одна пыльцевая клетка а соединяется с зачатковой клеткой А, другая – с а.
Результаты этих оплодотворений можно сделать показательными, обозначая связанные зачатковые и пыльцевые клетки в форме дроби, причем пыльцевые клетки помещаются над чертой, а зачатковые клетки – под ней. В данном случае получается:
А/А + A/а + a/A + a/а
У первого и четвертого члена зачатковые и пыльцевые клетки одинаковы, поэтому продукты их соединения константны, а именно А и а; у второго и третьего, наоборот, происходит вновь соединение обоих различающихся исходных признаков, поэтому выходящие из этого оплодотворения формы совершенно тождественны с гибридом, от которого они произошли. Следовательно, происходит повторная гибридизация. Этим объясняется то замечательное явление, что гибриды могут производить наряду с обеими основными формами также потомков подобных себе; A/a и a/A дают оба одинаковое соединение Аа, потому что для успеха оплодотворения, как было показано раньше, нет никакой разницы, какой из двух признаков принадлежит пыльцевой или зачатковой клетке. Поэтому
A/A + A/a + a/A + a/a = A + 2Aa + a
Такой характер принимает в среднем ход распределения форм при самооплодотворении гибридов, если в них соединяются два различающихся признака. Но в отдельных цветках и у отдельных растений могут происходить значительные отклонения в относительном количестве производимых ими отдельных форм ряда. Несмотря на то, что количества, в которых встречаются оба рода зачатковых клеток в завязи, являются равными только в среднем, вполне предоставляется случаю, каким из двух видов пыльцы произведено опыление каждой отдельной зачатковой клетки. Поэтому отдельные цифры неизбежно подвергаются колебаниям, и возможны даже крайние случаи, которые приводились раньше в опытах над формой семян и окраской белка. Истинные числовые отношения могут быть даны только средними величинами, полученными из суммы возможно большего числа отдельных случаев; чем больше их число, тем вернее устраняются случайности.
Ряды развития гибридов, в которых соединены двояко различающиеся признаки, содержат 9 различных форм среди [каждых] 16 индивидов, именно: АВ + Аb + аВ + ab + 2АВb + 2аВb + 2АаВ + 2Ааb + 4АаВb. Между различными признаками исходных растений А, а и В, b возможны 4 комбинации; поэтому и гибрид дает четыре соответствующие формы зачатковых и пыльцевых клеток АВ, Аb, аВ, ab, и каждая из них вступает в среднем 4 раза в оплодотворение, если в ряду содержится 16 индивидов. Поэтому в оплодотворении принимают участие:
