


The members of these three orders have features in common which have been evolved independently as essential adaptations to marine life. The most conspicuous of these is the streamlining of the animal by the production of a thick layer of blubber under the skin which not only smoothes out angularities but also provides an insulating layer against the low temperature of the water. In addition the limbs are reduced in all, the long bones of the fore-limbs are shortened and in the Cetacea and Sirenia the hind-limbs are lost altogether, while in the Pinnipedia these hind-limbs are much modified. The body too is elongated and roughly spindle-shaped in its proportions. These modifications deal with two problems connected with water, namely its low temperature and its greater density. There remains a third which is in some ways more serious, namely, that these animals being mammals are firmly committed to air breathing, possessing lungs, so that access to the air is essential and a means of preventing the entry of water into the trachea and lungs equally so. All therefore have nostrils which are normally closed and are opened only by voluntary muscles when the head is above the surface of the water. The provision of oxygen to maintain activities when the animal is submerged is made in different ways in the different groups. For example Cetaceans dive with full lungs, pinnipedes with empty ones, but the details of respiration are complex and will be dealt with later. Propulsion through the dense medium of water has also led to another convergent feature, the fusing of the digits by webbing, either thin or thick, thus forming a flipper or fin out of the normal mammalian hand or foot. There is also a tendency for the hair to be reduced in length although this is not universal in the groups. In Cetacea and Sirenia the body is almost naked, but the vibrissae or moustachial hairs are retained either as normal tactile organs as in the Sirenia or much reduced in the Cetacea. In neither of these groups does the hair contribute to insulation. Nor does it do so in the hair-seals and other pinnipedes where the hair is short and easily wetted so that the water comes into direct contact with the skin. Only in the fur-seals is the hair dense so that on immersion a layer of air is trapped among the hairs of the undercoat and direct water to skin contact is prevented. Here the hair is accessory to the blubber as an insulating structure.
We are now beginning to deal with features in which there are considerable and obvious divergences between the groups and these can best be described by noting them in the pinnipedes and then briefly contrasting them with what appears in the other two orders. Unlike the Cetacea and Sirenia which are entirely aquatic throughout life, the Pinnipedia have not lost all contact with the land. Some spend more time in the water than others, but all come to land (or ice) for breeding and for basking between feeding bouts and therefore have retained an ability to move on land. This is achieved by two different methods, one found in the true seals and the other in the fur-seals, sea-lions and walruses (Fig. 1). In these latter the hind-limbs are still capable of being directed forwards and acting as a foot, albeit on an extremely short leg which is buried in blubber down to the ankle. Thus with the associated flexure of the trunk in the lumbar region they can move rapidly over a land surface. The true seals, however, have hind-limbs which are directed backwards and can only trail on land. Movement is therefore much more laboured in a terrestrial habitat. A comparison between the limbs of a furseal or sea-lion and those of a true seal shows clearly the very considerable differences not only in structure and posture, but also in use.
The fore-limbs of a fur-seal are long and on land are capable of reflexion between the wrist and palmar surface. The forearm and wrist form a vertical prop as it were, while the hand, supported by the meta-carpal and digital bones, lies flat on the ground (Fig. 2a). The web between the digits is thin, in fact the whole of the distal part is much thinner and longer than in the true seals. This is associated with the much greater use which the fur-seals make of their fore-limbs in swimming. The claws are quite rudimentary and in some almost missing altogether. In the true seals the fore-arm is buried in blubber, and only the wrist and palmar surface with short digits protrude as short flaps (Fig. 1). The webbing cannot be distinguished as such for the digits are united by thick tissue so that the separate digital lines are not visible externally. The claws are strong and used for grooming the surface of the body. In the water these limbs are used principally for changing direction or for slow paddling, never for rapid movement, when they are held tightly pressed against the flanks. The digits still retain, as Backhouse has shown, an ability to flex with considerable power so that the animal can haul itself up over boulders and rocks and to some extent compensates for the loss of power of the hind-limbs. At any rate this is true for the northern true seals. In the antarctic species this ability if present is not used, so far as observations made by a number of observers appear to confirm (O’Gorman).
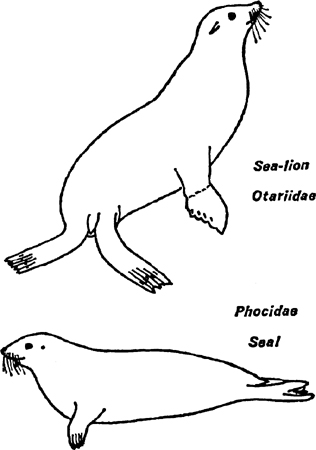
FIG. 1. External features of seal and sea-lion. Note the differences in the external ear, the longer fore-limb of the sea-lion and the way the hind-limbs of the sea-lion can turn forward compared with the trailing position in the seal. The sea-lion can raise the forepart of the body into an almost erect position.. The claws of the sea-lion are small and set back from the edge of the flippers. (See also Plate 1)
The hind-limbs of a fur-seal are also long but can be directed forwards for movement on land. They are also used on land during basking as fans. When fur-seals are hauled-out for breeding the bulls have to remain on their territories for a long time and they soon dry out. Their layer of blubber and the thick undercoat of fur, which keeps them warm in water, is now a disadvantage when the sun comes out. They then use the hind flippers as huge fans, turning the body on one side so that both flippers can fan the anterior part of the body and head which has less fur on it (Pl. 1). The claws here are usually absent altogether or quite rudimentary on the first and fifth digit but well developed in the middle three which can thus be used for grooming. In the true seals the hind-limbs are permanently directed backwards and form powerful sculling organs (Fig. 2). The palmar surfaces of the feet are turned inwards to face each other. The digits are united by thin and extensive webbing and the digits themselves are strong, the outer ones (the first and fifth) being much longer and thicker than the others. By alternate sweeps from side to side in a sculling movement these flippers are able to drive the seal through the water at great speed which has been estimated at as much as 12 to 15 m.p.h. Claws are retained on all the digits but their function is obscure. On land these hind flippers trail behind and take no part in locomotion which is achieved by a ‘humping’ motion. The body is flexed and the pelvic region advanced to give a forward push to the anterior part of the body, the stomach and the chest taking the main load as the animal moves forward. The fore-limbs are not used in this motion unless obstacles have to be surmounted when they assist in dragging the body upward, helped by the thrust of the pelvis. Thus in the Pinnipedia there are two distinct means of progression both on land and in water.
In the Cetacea the principal locomotor organ is the tail, flattened dorso-ventrally into a triangular fluke, and moved up and down in a sort of ‘crawl’ action. The hind-limbs are missing and the fore-limbs are well developed into flippers. Not only are the digits joined by thick webbing but the number of phalangeal bones is greatly increased so that the flipper becomes extremely flexible in its use as a propulsive organ and also in causing change in direction. The Sirenia are rather lethargic and movement is obtained by the action of the flattened tail and by the broad, short fore flippers. The latter are peculiar in having a great power of rotation so that their direction of action can vary over a wide angle. This is of considerable advantage when used as a paddle, the animal being able to turn almost in its own length, or when erect in the water, in little more than its own breadth.
A feature associated with streamlining is the reduction of the pinna or external ear flap. In the Pinnipedia all variations are found; among the fur-seals and sea-lions it is still fairly well developed but narrow (furled) and elongate, lying back alongside the head. For this reason they are often called the ‘eared’ seals. In the true seals it is much reduced and does not extend beyond the dried hair. When the hair is wet it can just be seen, if the light is right, as a circular rim round the earhole (Pl. 1). In the walrus it is equally inconspicuous. In both Cetaceans and Sirenia it is completely absent and in the former the earhole is plugged. (Only recently has the extraordinary hearing mechanism of whales and porpoises been demonstrated by Drs Fraser and Purves.)
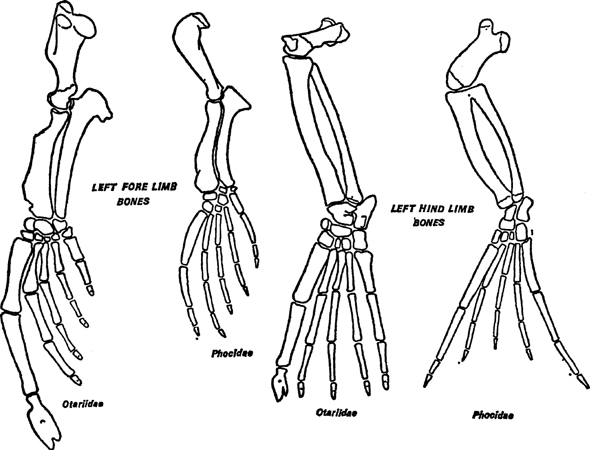
FIG. 2. Limb bones of seal and fur-seal: left, fore-limb skeletons; right, hind-limb skeletons. Note particularly the relatively small fore-limb of the seal (Phocidae) and the almost equal size of the ‘big’ and ‘little’ toes in the hind-limbs. In the Otariidae the ‘thumb’ and ‘big’ toe are much larger than the other digits, indicating the driving use made of both limbs. Notice too the very short thigh (femur) and upper arm (humerus) bones in both seal and fur-seal. For comparisons the hind-limbs of both are drawn to the same length and the fore-limbs to the same scale. c. x
The normal resting closed position of the nostrils has already been mentioned as common to all three groups. It remains to point out that the pinnipedes retain a ‘rhinarium’ external to the nostrils. This is the equivalent of the ‘wet nose’ of the dog and must constitute an important sense organ for the reception of chemical stimuli (scent). That it has to operate in water is no drawback because the molecules of the ‘scent’ must be dissolved before they can stimulate a receptor nerve ending, for which reason the rhinarium of land mammals is always kept moist with secreted mucus. In the pinnipedes it is possibly the sense organ which locates prey, such as shoals of fish at a distance.
The teeth of pinnipedes are greatly modified from the normal mammalian pattern. The differentiation into incisors, canines and molars is still recognisable, but they do not differ so much from each other as in other (land) mammals, consisting basically of single pegs of varying lengths. The incisors are small and degenerate as shown by the variation in their number. (It is not unusual to find two on one side and three on the other.) The canines are massive cones, used for offensive purposes in the males of some species but basically the prey-catching tooth as in land carnivores. The molars are no longer grinders or mashers but consist of a single major cusp or spike with one or more smaller cusps on each side in line with the length of the jaw. In this way there is formed a long line of pointed cusps of varying heights admirably adapted to the retention of struggling prey. The only exceptions are the walrus and the bearded seal. The walrus feeds on large bivalve molluscs, like clams and mussels, which are crushed between stub-like molars. The bearded seal is not predominantly a fish feeder, shrimps, crabs, holothurians, clams, whelks, snails and octopus forming the bulk of its food and the molars are usually much worn and not sharply pointed. In some species of seal the young do have milk teeth for a short time. Usually there is only one set of functional teeth, the milk dentition being resorbed while the foetus is still in utero. Cetacea either have no teeth and feed by a filter mechanism (whale-bone whales) or have a single row of simple cusps (the toothed whales, porpoises and dolphins), all alike, which are far more numerous than in any other mammal. This is a high specialisation for fish-eating, much greater than the pinnipede condition. The Sirenia have flattened grinders of degenerate form since the seaweeds on which they feed hardly require munching.
The skulls of most pinnipedes show traces of slow and incomplete ossification. This is particularly true of the region on a level with the eyes so that the front part can be easily detached from the hinder brainbox in even old animals. The fur-seals and seals differ considerably in the general appearance of their skulls thus lending support to the view that they are derived from different sections of the land carnivores (Fig. 3). The walrus is again an exception since there is massive ossification to provide support for the huge tusks. Cetacea have evolved quite differently since many of the cranial bones contain spaces filled with air or occasionally with oil. The Sirenia have massive skulls although the bone itself is not dense.
We now come to the respiratory modifications and it is impossible to separate these from peculiarities of the blood system since the oxygen required by the tissues is transported by the haemoglobin in the red cells of the blood. The modifications of the nostrils have already been mentioned, but there are others equally significant. Many pinnipedes have cartilaginous rings in the trachea which are incomplete on the upper side but some, and both of our British species are among them, have complete rings which thus prevent any collapse of the trachea when the seal is under pressure in diving. These rings are continued into the bronchi and bronchioles and cartilage continues to be found in the connective tissue of the lungs lying between the respiratory lobules. In addition there are, in the bronchi, valves of muscle and connective tissue which are able to form air-locks in the lungs and so prevent the residual air in the larger (and non-respiratory) tubes being forced under pressure into the respiratory alveoli. This appears to be a device to prevent nitrogen, which forms four-fifths of the air, being absorbed under pressure into the blood stream. If it were so absorbed, on return to the surface it would come out of solution in the blood under the reduced pressure to form gas bubbles in the smaller blood vessels and so cause ‘bends’ which can easily prove fatal, as it does when it occurs in man when diving.
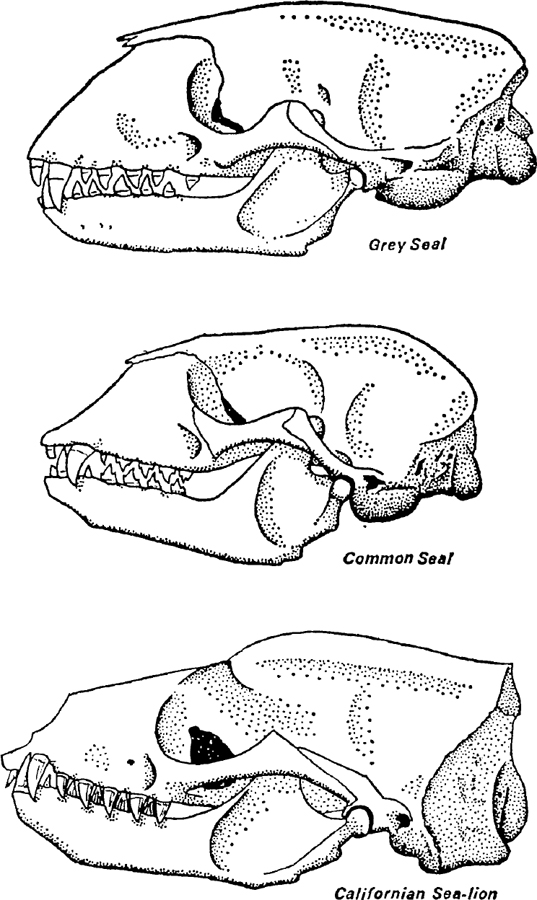
FIG. 3. Skulls of seals and sea-lion. All the skulls are of adult males. For comparisons the grey seal skull is drawn to the same length as that of the Californian sea-lion and the common seal skull to the same scale as that of the grey seal. Note the general similarities, particularly of the dentition, but also the differences which are most marked in the region behind the articulation of the lower jaw. c. x
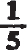
The lungs themselves are not abnormally large and all pinnipedes exhale before or on diving so that there is literally little or no oxygen in the lungs to provide for tissue respiration during the activities of swimming and catching prey below the surface. The thorax, which also contains the heart and great vessels besides the lungs, is more elongate than normal in mammals and the diaphragm, which separates its cavity from that of the abdomen, is set much more obliquely, its upper attachment to the body wall being set farther back than normal and the sternal support of the lower margin is shorter than usual (Fig. 4). This means that the cavity can be more completely compressed and a greater proportion of the air in the lungs exhaled than in normal land carnivores and other mammals. The small residium is driven into the nonrespiratory trachea and bronchi. When the seal returns to the surface breathing recommences and a series of deep inhalations and expirations takes place. From my own observations on a southern elephant seal the number of such breaths is roughly proportional to the length of time that the nostrils have been closed. Even when on land and hauled-out seals will continue to remain with closed nostrils for considerable periods separated by series of breathings.
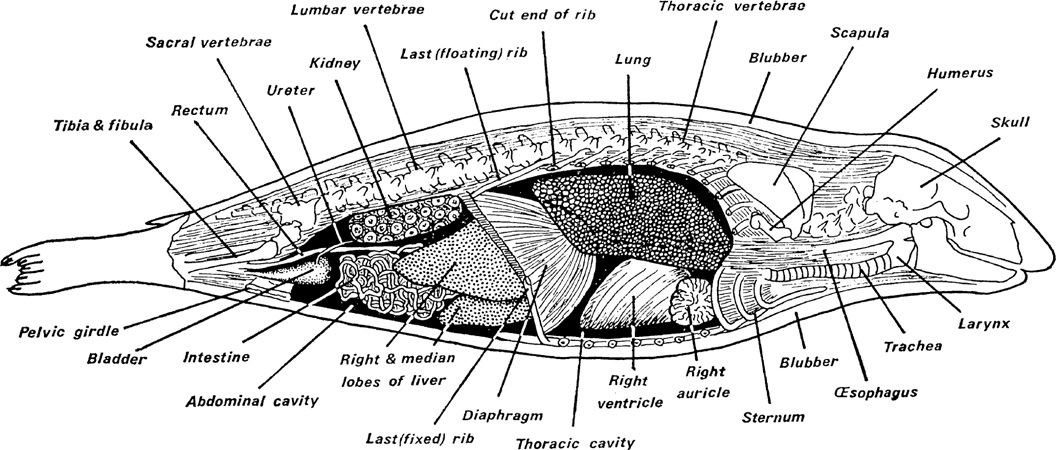
FIG. 4. Side view of the pinnipede body, to show the principal organs. The right side of the body is diagram-matically opened by removing the skin, blubber and outer muscle layers. The right fore-limb has also been removed as far as the ‘elbow’. The genitalia are omitted. The oesophagus is indicated as passing above the heart and between the right and left lungs.
By way of contrast Cetaceans dive with full lungs and all the modifications are towards the prevention of collapse and the transmission of the external pressure to the air contained in the lungs; another ‘anti-bends’ device.
In the pinnipedes we are still left with the puzzle of how they obtain and maintain sufficient oxygen for their activities below the surface, and we must turn to the blood system for further information.
First it must be clear that if there is little or no air in the lungs there is no profit in circulating the blood from the lungs for there is no oxygen to pass on to the active tissues. It is therefore not altogether surprising to find that seals exhibit a phenomenon known as ‘bradycardia’. This is a reduction in the heart beat both in the number per minute and in the strength of the beat. In fact it is reduced to little more than an occasional flutter by which some blood is circulated along the carotid artery to the brain. This has been shown to arise almost immediately the seal has dived, and in this it differs from the bradycardia of Cetaceans in which the rate and strength of the heart beat is gradually reduced to a low level. This difference must be associated with the difference of lung contents, the gradual bradycardia of Cetaceans keeping pace with the gradual exhaustion of the oxygen in the lungs (Fig. 5).
To prevent the ‘used’ blood from the tissues of the body being circulated even to a very minor degree in the pinnipedes, they have evolved a powerful sphincter muscle which closes the huge venous blood vessel leading to the heart and which draws blood from the hinder part of the body, the viscera and liver. This large blood vessel (posterior vena cava) is disproportionately large (usually double and enlarged) and so can act as a reservoir for the non-circulating blood. In addition there is a large vein lying below and up the sides of the spinal cord (extradural vein) which is enormously enlarged in pinnipedes. From it only a little blood can find its way back to the heart in the front region. Elsewhere this vein is connected by special large veins both directly to the posterior vena cava (at the hinder end where it is double) and indirectly round the kidneys in huge blood sinuses (Fig. 6). All these peculiarities increase the storage capacity of the venous system when bradycardia is in action. Some idea of the size of the veins will be conveyed by saying that in the grey seal the posterior vena cava in its posterior part is ‘nearly as thick as your wrist’ and King (1964) refers to their size in the walrus ‘by the often quoted reference that they can be “pulled on like a pair of trousers”’.
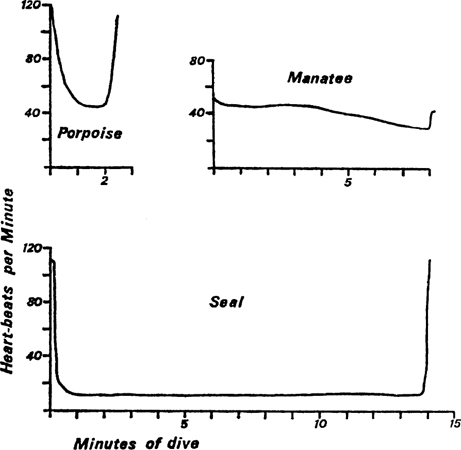
FIG. 5. Bradycardia in seal, porpoise and manatee. These are representatives of the three groups of truly marine mammals, the manatee, a surface feeder on seaweeds, being the least modified for diving. The seal immediately responds to diving by reducing heart activity, having expelled all the air from the lungs, while the porpoise, retaining air in the lungs, reduces the heart-beat rate only slowly. (Redrawn from Irving, Scholander and Grinell, 1941)
But all these modifications tend to show that the blood is not the continuing source of oxygen during active diving. This is confirmed by two other facts; firstly the red blood cell count (the number of red blood cells per unit volume) is nothing out of the way, about 5–6 million (cf. human 4–5 million), and secondly that the amount of haemoglobin in a unit volume is not very high either, about 1.2 compared to the standard in man of 1.0. If this is all true, then how do seals manage to respire in their tissues during diving?
Part of the answer lies in another pigment known as myoglobin because it is present in muscle. Here we find an enormous difference from the normal and we hardly need figures to show it. Myoglobin is also coloured though not so deeply as haemoglobin and it gives the normal pink colour to muscle meat. In the Pinnipedia the muscle is almost black in colour, certainly very deep red. Those who remember the whale meat which was available after the second world war will recognise that Cetaceans too have a very high myoglobin content. To both of these groups then part of the answer is the ability to store oxygen attached to the myoglobin on the site where it is required for the respiration of the active muscle cells.
There is also evidence that these animals can run into oxygen debt, particularly in respect of metabolising the waste products, which are normally toxic if allowed to accumulate without treatment. This adaptation is not so extraordinary as it sounds, for it is known that, in humans, when slow starvation is prolonged and the organism begins to live on its muscle protein (autolysis of the muscle) a level of waste nitrogen products can build up to many times the normally lethal concentration. In these diving mammals this ability has become normal rather than a pathological occurrence. On return to the surface the repeated inhalations and exhalations rapidly restore the oxygen balance.
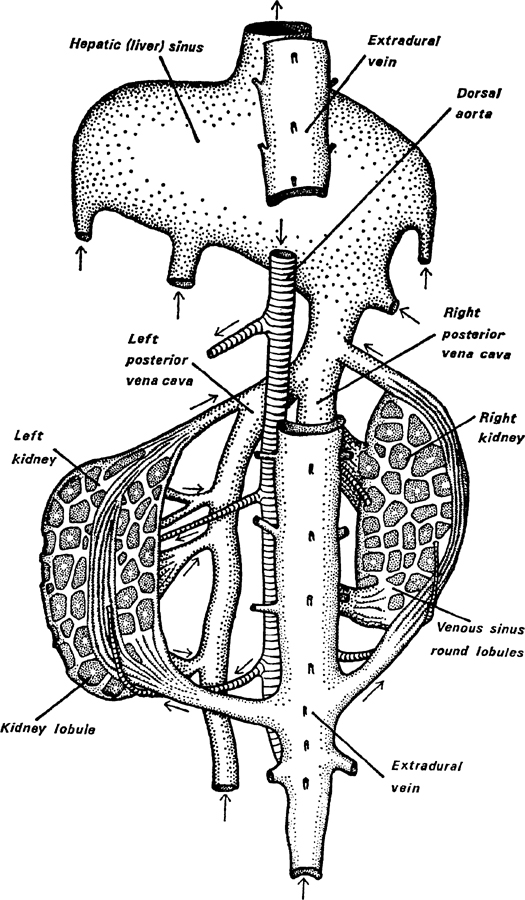
FIG. 6. Kidneys and posterior vena cava, showing the extra storage space in the blood sinuses. This is a view from above, the vertebral column and nerve cord being omitted. Part of the extradural vein is also omitted so as to allow the junction of the right and left branches of the posterior vena cava to be seen, with its continuation into the huge hepatic (liver) sinus. Arrows indicate the direction of the blood flow. (Redrawn and somewhat modified from Harrison and Tomlinson, 1956) X
So atypic of mammals is the environment of the pinnipedes that speculation as to the value of their principal sense organs must come to mind. Not a great deal has been found out but Harrison, working on captive common seals, has contributed some useful basic information. He finds that the eyes are well adapted for night vision or for murky water but this is associated with short-sightedness. The Weddell seal, however, avoids this myopia by having a slit pupil. Their effectiveness must therefore be strictly limited particularly in those species which live and feed through the polar winter. Auditory and touch stimuli are well received, the latter being particularly relevant to the well-developed vibrissae on the muzzle which have very large nerves running from them to the brain. Water being an incompressible medium is excellent for the transmission of sound waves, so that sensitivity to auditory stimuli is to be expected. My own impression is that grey seals are more sensitive to sounds in the higher frequency range than those in the lower. Phoca and Zalopus both have good hearing in the upper range. It is possible that they can hear sounds which, to man, are ultrasonic but this has not been proved. Recent work has shown that many marine organisms make use of these higher frequencies and sensitivity to them may well be an advantage to carnivores in search of food. Harrison also found that the common seals have an excellent sense of orientation. This is obviously of great benefit to an animal making use of a three dimensional medium such as the sea. Anyone who has done any flying will know that man, although he has the necessary sense organs, has to learn how to use and interpret the stimuli received.