


Полная версия
Руководство по спортивной медицине

Руководство по спортивной медицине
УСЛОВНЫЕ СОКРАЩЕНИЯ
АД – артериальное давление
АДФ – аденозиндифосфорная кислота
АКТГ – адренокортикотропный гормон
АМо – амплитуда моды
АМФ – аденозинмонофосфорная кислота
АТФ – аденозинтрифосфорная кислота
БАД – биологически активная добавка в/в – внутривенно
ВК – врачебный контроль
в/м – внутримышечно
ВНИИФКС – Всероссийский научно-исследовательский институт физической культуры и спорта
ВНС – вегетативная нервная система
ВПН – врачебно-педагогические наблюдения
ДМТ – дефицит массы тела
ДНК – дезоксирибонуклеиновая кислота
ДНЛЖ – давление наполнения левого желудочка
ДСН – дополнительная стандартная нагрузка
ЕИ – единица интенсивности
ЖЕЛ – жизненная емкость легких
ЖИ – жизненный индекс
ЖКТ – желудочно-кишечный тракт
ИВТ – исходный вегетативный тонус
ИГСТ – индекс Гарвардского степ-теста
ИК – индекс Кердо
ИМАО – ингибиторы моноаминоксидазы
ИМТ – индекс массы тела
ИН – индекс напряженности
ИР – индекс Руфье
ИС – индекс скелии
ИФС – интенсивность функционирования структур
ИЭ – индекс Эрисмана
КоА – коэнзим А
КПД – коэффициент полезного действия
Кр – креатин
КрФ – креатининфосфат
КФ – креатинфосфат
КЭК – коэффициент экономизации кровообращения
ЛПУ – лечебно-профилактическое учреждение
ЛФК – лечебная физическая культура
МЕ – моторная единица
МК МОК – Медицинская комиссия Международного олимпийского комитета
МО – минутный объем
Мо – мода
МПЗ – моторная плотность занятия
МПК – максимальное потребление кислорода
НАДН – никотинамидадениндинуклеотид
ОГК – окружность грудной клетки
ОМЦ – овариально-менструальный цикл
ОПСС – общее периферическое сопротивление сосудов
ПДГ – пируватдегидрогеназа
ПОЛ – перекисное окисление липидов
ПП – показатель Пинье
ППБЦ – продукты повышенной биологической ценности
РНК – рибонуклеиновая кислота
рРНК – рибосомная РНК
РФ – регулятор фосфорилирования
РЭГ – реоэнцефалография
СДГ – сукцинатдегидрогеназа
СЖК – свободные жирные кислоты
СИ – сердечный индекс
СилИ – силовой индекс
СПР – саркоплазматический ретикулум
ССС – сердечно-сосудистая система
СЭС – санитарно-эпидемиологическая станция
ТМТ – тощая масса тела
УВЧ – ультравысокая частота
УЗИ – ультразвуковое исследование
УМО – углубленное медицинское обследование
УО – ударный объем
ФК – функциональный класс
ФР – физическая работоспособность
цАМФ – циклический аденозинмонофосфат
ЦНС – центральная нервная система
ЧСС – частота сердечных сокращений
ЭКГ – электрокардиограмма
ЭхоКГ – эхокардиография
L – рост
P – вес
PWC – (Physical Working Capacity) физическая работоспособность
ПРЕДИСЛОВИЕ
Спортивная медицина как область медицинских и спортивных знаний в настоящее время стремительно развивается. Повышение внимания к этой области связано, в первую очередь, с беспокойством о стратегическом потенциале – здоровье народонаселения страны. Не секрет, что за последние два десятилетия уровень физической подготовленности, как в целом населения России, так и подрастающих поколений, существенно снизился. Поэтому перед спортивной медициной стоят важнейшие задачи – восстановление и поддержание здоровья целой нации.
Известно, что двигаться полезно всем: и грудному ребенку, и олимпийскому чемпиону. Весь вопрос в том, кому сколько?
Очевидно, что нагрузка должна быть дифференцированной и зависеть от многих факторов: пола, возраста, состояния здоровья, тренированности, окружающих условий и т. д. В зависимости от этих особенностей нагрузку необходимо сделать эффективной и безопасной. Кроме того, необходимо иметь знания по физиологии, биохимии, морфологии, чтобы на современном уровне оценить влияние занятий физкультурой и спортом на организм человека и таким образом выйти на управление процессами тренировок спортсменов и занятий оздоровительной физкультурой. В этом, на наш взгляд, и состоит идея создания настоящего руководства по спортивной медицине.
Считаем полезным ознакомить читателей с нормативно-правовым обеспечением в области спортивной медицины. Это позволит разъяснить основные положения действующих нормативных документов и ответить на вопросы терминологии и понятийного аппарата в спортивной медицине; объяснить такие права и обязанности специалистов, как устройство на работу, процедуры аттестации, лицензирования, аккредитации, повышения квалификации.
Возможно, данное руководство не является совершенным, могут встретиться дискуссионные моменты. Тогда авторы с благодарностью примут конструктивные замечания.
Профессор В. А. МаргазинЧАСТЬ I. ТЕОРЕТИЧЕСКИЕ ОСНОВЫ СПОРТИВНОЙ МЕДИЦИНЫ
ГЛАВА 1.
ФИЗИОЛОГИЯ ДЕЯТЕЛЬНОСТИ И ОСНОВЫ БИОЭНЕРГЕТИКИ МЫШЦ
1.1. Структура и свойства скелетных мышц.
Механизм мышечного сокращения
Особенностью мышц является их способность избирательно трансформировать химическую энергию в механическую. Последняя проявляется в виде силы или движения. Механическая энергия мышц затрачивается на потребности вегетативных функций внутри организма или на выполнение функций, связанных с взаимодействием организма и внешней среды. В первом случае используется энергия гладких мышц, во втором случае – энергия поперечнополосатой мускулатуры.
Скелетная мышца состоит из множества функциональных единиц – мышечных волокон или клеток. Они имеют цилиндрическую форму и расположены параллельно друг другу. Это многоядерные клетки 0,01 – 0,1 мм в диаметре, достигающие нескольких сантиметров в длину. К каждому мышечному волокну через специальную зону контакта – синапс – присоединено окончание нервного волокна. В свою очередь, каждое мышечное волокно состоит из 1000 – 2000 параллельно расположенных субъединиц (диаметром около 1 мкм), называемых миофибриллами, которые включают в себя повторяющиеся в продольном направлении блоки – саркомеры. Оба конца мышечного волокна образованы соединительной тканью, посредством которой волокно прикрепляется к костям скелета. В целой мышце эта соединительная ткань образует сухожилие, или апоневроз. Нормальная функция мышцы зависит от тех влияний, которые она получает из нервных центров благодаря нервным волокнам. Функциональное единство поперечнополосатой скелетной мышцы обеспечивается, таким образом, нервным волокном и совокупностью иннервируемых им мышечных волокон. Каждое моторное нервное волокно (аксон), являющееся отростком двигательной клетки передних рогов спинного мозга, иннервирует не одно, а целую группу мышечных волокон. Такая группа получила название моторной (или двигательной) единицы (МЕ). Количество мышечных волокон, входящих в ее состав, варьируется от 10 до 3000 ME. Наименьшее число волокон содержится в быстрых мышцах, обеспечивающих точные тонкие движения. Так, в МЕ глазных мышц и мышц пальцев руки содержится по 10 – 25 мышечных волокон, причем каждое из них получает иннервацию от нескольких нервных волокон. С другой стороны, мышцы, обеспечивающие поддержание позы, состоят из МЕ, имеющих в своем составе 2000 – 3000 волокон.
Виды сокращений. В экспериментах над животными мышечное сокращение обычно вызывают электрическим раздражением. Раздражитель может состоять из одного изолированного электрического стимула (длительностью, например, 1 мc) или содержать серию электрических стимулов (повторяющаяся стимуляция).
У скелетных мышц выделяют соответственно одиночное сокращение и суммированное (тетанус).
Одиночное сокращение возникает в ответ на одиночный пороговый стимул. В нем выделяют три фазы: латентный период, фаза укорочения и фаза расслабления. Во время латентного периода не регистрируется никаких механических феноменов, по его прошествии наступает фаза укорочения, во время которой быстро нарастает напряжение мышцы. Фаза расслабления продолжается примерно в 2 раза дольше, чем фаза укорочения. Для одной и той же мышцы время сокращения увеличивается при снижении температуры или при утомлении мышцы. Для мышц с очень высокой быстротой сокращения, в частности для глазодвигательных мышц, время сокращения составляет 7 – 10 мc; для мышц конечностей это время длится от 25 до 40 мc. Для камбаловидной мышцы (M. soleus), играющей важную роль в сохранении позы и имеющей низкую быстроту сокращения, это время достигает 90 – 120 мc. Мышцы с высокой быстротой сокращения образуют бледные волокна, у мышц с малой быстротой сокращения волокна красные, богатые миоглобином. Мышцы-сгибатели принадлежат к первому типу, разгибатели, играющие важную роль в поддержании позы, – ко второму.
Сокращение, как и возбуждение, распространяется вдоль мышечных волокон. Скорость прохождения волн возбуждения и сокращения одинакова. Например, для двуглавой мышцы плеча скорость возбуждения составляет 3,5 – 5 м/с. Ввиду относительно большой длительности сокращения наступает момент, когда вся мышца сокращена, так как последние ее участки, до которых доходит волна возбуждения, уже сократились, а первые еще не расслабились. Этот момент соответствует моменту максимального укорочения.
Величина одиночного сокращения прямо пропорциональна силе раздражения. Однако при достижении определенной силы раздражения величина сокращения перестает расти, несмотря на дальнейшее повышение силы раздражения. Это объясняется тем, что каждое волокно обладает разной возбудимостью и реагирует по закону «все или ничего». При пороговой силе реагируют наиболее возбудимые волокна. Чем сильнее раздражитель, тем больше волокон возбуждается. При максимальном сокращении возбуждаются все волокна мышцы.
В естественных условиях в организме скелетная мышца получает обычно из нервной системы не одиночные раздражения, а ряд быстро следующих друг за другом нервных импульсов. Под влиянием ритмических раздражений наступает длительное и сильное укорочение мышцы, которое называется тетаническим сокращением, или тетанусом. То, что тетанус возникает в результате действия серии одиночных возбуждений, доказывает регистрация потенциалов действия в тетанически сокращенной мышце. Так, при произвольных движениях руки у человека число потенциалов действия достигает 50 – 70 в секунду.
Тетанические сокращения представляют собой результат суммации одиночных сокращений. Для искусственного воспроизведения тетануса на мышцу действуют серией раздражителей, следующих друг за другом. Если поступает несколько раздражений с интервалом большим, чем фаза укорочения, то возникает явление, которое называется зубчатым тетанусом. При частоте следования раздражений, которая меньше фазы укорочения или равна ей, возникает гладкий тетанус. Амплитуда гладкого тетануса зависит от частоты раздражения. Если каждый последующий раздражитель поступает в фазу экзальтации (повышенной возбудимости), то ответ мышцы будет достаточно большим. Если раздражитель поступает в период сниженной возбудимости (относительная рефрактерная фаза), то ответ мышцы будет много меньше. Такая зависимость амплитуды ответа от частоты раздражения получила название оптимума и пессимума частоты раздражения. Например, á-мотонейрон может посылать к мышце импульсы с частотой 20 имп/с, 40 имп/с, 50 имп/с. В зависимости от частоты величина сократительного ответа будет различной. Это один из способов регуляции силы мышечных сокращений. В реальных условиях все сокращения скелетных мышц возникают в ответ на периодическую стимуляцию и являются тетаническими. Сравнение отдельного мышечного сокращения с тетаническим показывает, что напряжение, развивающееся при тетанусе, выше максимального напряжения, регистрируемого во время одиночного; разница часто достигает соотношения 4: 1.
Для скелетной мышцы характерен еще один вид активности – контрактура. После прекращения тетанического раздражения мышечные волокна расслабляются вначале не полностью – их исходная длина восстанавливается лишь по истечении некоторого времени. Это явление называется послететанической (остаточной) контрактурой. Природа контрактур заключается в остаточной длительной деполяризации мышечной мембраны. В целостном организме контрактура возникает при условиях патологии и проявляется в длительном слитом сокращении мышцы.
Режимы сокращения скелетных мышц. Для скелетных мышц характерны два основных режима сокращения: изометрический и изотонический. Изометрический режим проявляется тогда, когда в мышце во время ее активности нарастает напряжение (генерируется сила), но из-за того, что оба конца мышцы фиксированы, например мышца пытается поднять непосильный груз, она не укорачивается. Изотонический режим проявляется, когда мышца первоначально развивает напряжение, способное выполнить работу, например поднять данный груз. Потом мышца укорачивается – меняет свою длину, сохраняя напряжение, равное весу поднятого груза. Так как изотоническое сокращение не является исключительно изотоническим (элементы изометрического сокращения имеют место в самом начале сокращения мышцы), как и изометрическое тоже не является полностью изометрическим (элементы смещения все-таки есть), то предложено употреблять термин ауксотоническое сокращение, т. е. смешанное по характеру.
В реальной практике оба режима могут переходить из одного в другой. Так, когда мышечная стенка полого органа начинает сокращаться, а орган содержит жидкость и выход из него закрыт сфинктером (сердце, мочевой пузырь), то возникает ситуация изометрического режима: давление внутри полого органа растет, а размеры его не могут измениться, так как жидкость не сжимается. На определенной величине давления сфинктер открывается и мышца переходит в изотонический режим сокращения на время изгнания жидкости из полости органа, когда напряжение сохраняется на постоянном уровне.
Механизм мышечного сокращения. Для того чтобы понять сущность процесса сокращения мышечного волокна, необходимо вспомнить, что каждое мышечное волокно состоит из миофибрилл, которые включают в себя повторяющиеся в продольном направлении блоки – саркомеры, отделенные друг от друга так называемыми Z-пластинками (сеть из переплетающихся белковых молекул). Сверху миофибриллы покрыты мембраной мышечного волокна – плазмалеммой, толщина которой порядка 10 нм. Функциональной единицей миофибриллы поперечнополосатой мышцы является саркомер. В обоих направлениях от Z-пластинки тянутся тонкие нити (филаменты), состоящие из белка актина. Они переплетаются с толстыми нитями белка миозина. Миозиновые нити образуют наиболее оптически плотную часть саркомера — А-диск. Светлый участок в середине А-диска называют зоной Н. Здесь находятся только миозиновые нити и нет актиновых. В середине Н-зоны располагается М-линия, в которой находятся ферменты (например, креатиназа), играющие важную роль в энергетическом метаболизме. На участках взаимного перекрытия каждая миозиновая нить окружена шестью актиновыми, а каждая актиновая – тремя миозиновыми. На миозиновых нитях обнаружены мелкие выступы, называемые поперечными мостиками. Они расположены вдоль миозиновых нитей в виде 2-нитчатой спирали. Во время сокращения эти мостики смыкаются с актиновыми нитями. Актиновая нить образована двумя спиральными тяжами из глобулярных молекул актина (G-актин), закрученными один вокруг другого. В продольных бороздках актиновой спирали находятся нитевидные молекулы белка тропомиозина. Это палочкообразный фибриллярный белок, палочки соединяются конец в конец и образуют два тяжа, закрученных в виде спирали вокруг актина. К каждой молекуле тропомиозина прикреплен комплекс молекул глобулярных белков под общим названием тропонин.
Молекула миозина состоит из двух частей: длинного палочкообразного участка, который подразделяют на шейку и хвост, и глобулярного участка, представленного двумя одинаковыми головками. Диаметр этого участка молекулы составляет 4 нм, длина – 20 нм. Если молекулу миозина обработать протеолитическим ферментом трипсином, то она распадется на две части: легкий меромиозин и тяжелый меромиозин. Последний, в основном образованный головкой и шейкой молекулы, обладает ферментативной и актинсвязывающей активностью. Миозиновые нити формируются объединением отдельных молекул. Хвосты молекул ориентированы к середине, а головки направлены в противоположные стороны и образуют выступы на нити. Молекулы миозина расположены в миозиновой нити так, что головки равномерно распределены по всей длине (кроме небольшого участка в середине).
При наблюдении в микроскоп было обнаружено, что при сокращении ширина А-дисков не изменяется, тогда как I-диски (изотропные) и зоны Н становятся более узкими, т. е. изменяется ширина взаимного перекрытия актиновых и миозиновых нитей. Исходя из этого, A. F. Huxley в 1954 г. предложил для объяснения механизма мышечного сокращения теорию скольжения нитей (рис. 1).
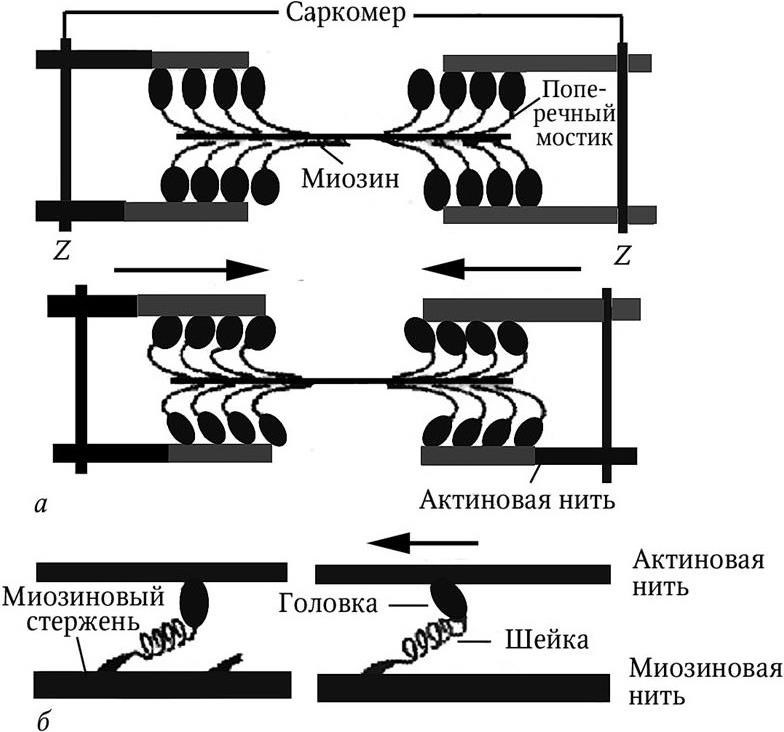
Рис. 1. Модель механизма сокращения – миозиновая нить с поперечными мостиками, прикрепленными к актиновым нитям:
а – схема сокращения мышечного волокна в пределах саркомера; Z – пластинка;
б —схема работы поперечного мостика (единицы соединения актина и миозина)
Согласно теории, укорочение саркомера происходит благодаря активному скольжению тонких актиновых нитей относительно толстых миозиновых в пределах саркомера. Длина самих нитей при этом не изменяется. Во время сокращения каждая головка миозина или поперечный мостик могут связывать миозиновую нить с актиновой. Наклоны головок создают объединенное усилие, и происходит «гребок», продвигающий актиновую нить к середине саркомера. Биполярная организация молекул миозина обеспечивает возможность скольжения актиновых нитей в противоположном направлении в обеих половинах саркомера.
Механизм этого процесса может быть объяснен за счет электромеханического сопряжения, т. е. передачи сигнала возбуждения с мембраны на миофибриллы. Ключевую роль при этом играют ионы Са2+. В расслабленном состоянии, т. е. при низкой концентрации ионов Са2+, регуляторные белки – тропомиозин и тропонин C – блокируют прикрепление поперечных мостиков миозина к актиновым нитям. Возбуждение вызывает высвобождение ионов Са2+ из саркоплазматической сети. Это ведет к тому, что концентрация Ca при возбуждении внутри волокна увеличивается и ионы Са2+ соединяются с молекулой тропонина. В результате происходит смещение молекулы тропомиозина, что обеспечивает возможность прикрепления миозиновых поперечных мостиков к актиновым нитям. Это конформационное изменение инициирует ряд процессов, приводящих к сокращению мышцы. Следовательно, в покое белки тропонин и тропомиозин препятствуют соединению мостиков миозина с актином. Активация Са2+ ведет к инактивации регуляторных белков, и происходит присоединение мостиков.
Следующий вопрос состоит в том, как мышца преобразует химическую энергию в механическую. Сами сократительные белки – актин и миозин не обладают аденозинтрифосфатазной (АТФазной) активностью. Однако, связавшись с актином, миозиновая головка поперечного мостика в присутствии ионов Мg2+ приобретает активность АТФазы и катализирует расщепление аденозинтрифосфорной кислоты (АТФ). Молекула АТФ связывается с активным АТФазным центром головки миозина и отделяет ее от актиновой нити. Гидролиз сопровождается конформационными изменениями головки молекулы миозина, переводя ее в высокоэнергетическое состояние.
Циклическая активность поперечных мостиков (ритмическое прикрепление и отсоединение), которая обеспечивает мышечное сокращение, возможна до тех пор, пока продолжается гидролиз АТФ, т. е. пока происходит активация АТФазы и существует достаточная концентрация ионов Са2+. В нормальных условиях расслабление мышцы происходит за счет того, что ионы Са2+, выделившиеся при возбуждении, начинают активно поглощаться саркоплазматическим ретикулумом (СПР). Уровень Са2+ в саркоплазме снижается, и тропомиозин начинает препятствовать присоединению поперечных мостиков. Мышца остается расслабленной до тех пор, пока вторично не произойдет следующая деполяризация мембраны.
1.2. Энергетика мышечной работы
Мышечную массу туловища составляют поперечнополосатые мышцы. Они являются самой объемной тканью тела человека, достигая у мужчин 50 % массы тела. Их деятельность контролируется сенсорными участками коры головного мозга и подкорковых центров движения.
Реализация движений и локомоций является основной функцией поперечнополосатых мышц. Кроме того, они являются самым крупным генератором биологической энергии в силу своей массы и выполняют функцию «второго сердца», способствуя возврату венозной крови в правое сердце, пропульсивно воздействуя на стенку периферических вен.
Все это в еще большей степени подчеркивает значимость движений для жизнедеятельности организма. Поперечнополосатые мышцы подвержены возрастной инволюции – устариков доля мышечной ткани уменьшается до 27 %, снижается тонус мышц и резко падает объем двигательной активности. С этим связано уменьшение роли мышц как генератора энергии и снижение выполняемой роли «второго сердца», что приводит к отекам, застоямит.д.
Коэффициент полезного действия (КПД) мышечной работы очень велик – 37 – 65 %. Энергия мышечного сокращения расходуется на изменение пространственного взаимоотношения сократительных белков.
Мышечная клетка. Наиболее значимыми белковыми фракциями мышечной клетки являются:
– белки миогеновой группы – в основном ферменты гликолиза и миоальбумин;
– миоглобин – красный хромопротеид, «родной брат» гемоглобина, определяющий красный цвет мышечной ткани; он содержит в своем составе свободный, легко окисляемый атом железа, связывает и транспортирует кислород в пределах цитоплазмы клетки;
– глобулины – ферменты и запасные белки, способные при тренировке преобразовываться в сократительные белки миофибрилл;
– миофибриллярные белки: миозин и актин, ферменты и др.;
– ядерные белки – нуклеопротеиды.
Наиболее значимыми из энергетических фракций являются такие водорастворимые азотистые соединения, как АТФ (0,25 – 0,4 % от сухого остатка клетки) и креатининфосфат (КрФ) – 0,4 – 1,0 % от сухого остатка клетки.
К важнейшим безазотистым энергетическим фракциям относится гликоген, который может составлять до 2 % сухого остатка клетки. Он может находиться в свободном и связанном состоянии, причем физическая тренировка значительно увеличивает количество свободного гликогена, что существенно повышает сократительную способность миофибрилл. Кроме этого, в механизме мышечного сокращения играют роль жиры, холестерин и минеральные соли.
Из других включений в цитоплазму клетки обращают на себя внимание митохондрии (митохондриальные кристы). Молекулярные биологи считают, что в них происходит аэробный синтез АТФ и синтез белков. В мембранах митохондриальных крист очень высок электрохимический потенциал ионов водорода, что позволяет рассматривать их как дополнительный источник внутриклеточной энергии, обеспечивающий эффективность перехода электролитов через клеточную мембрану.
Располагающиеся на клеточной мембране миоцита разноименно заряженные ионы металлов (калия, кальция, магния, натрия, кремния и др.) определяют работу «ионных мембранных насосов».
Мышечные волокна. Мышечные клетки структурно организованы в мышечные волокна. Общеизвестны два вида мышечных волокон (о них мы упоминали выше). Это белые мышечные волокна, называемые еще «быстрыми», и красные мышечные волокна – «медленные».
Белые волокна первыми откликаются на команду к деятельности со стороны центральной нервной системы (ЦНС), играя роль своеобразного стартера. В них преимущественно развиты анаэробные процессы ресинтеза АТФ, основным способом энергообеспечения является анаэробный гликолиз, который протекает в них с более высокой скоростью. Также белые волокна характеризуются относительно низким количеством митохондрий и миоглобина, высокой активностью миозиновой трифосфатазы, высокой буферной емкостью и хорошо развитым СПР. Они приспособлены к мощной, взрывной работе в короткий промежуток времени.
Красные мышечные волокна включаются в работу позднее белых, когда к тому возникнут определенные условия метаболизма. В этих волокнах низкая скорость гликолиза, но высокая интенсивность тканевого дыхания, высокое содержание миоглобина и митохондрий, низкая активность миозиновой АТФазы, низкая буферная емкость, значительно хуже развит СПР. Красные волокна предназначены для выполнения работы легкой либо умеренной мощности, но в течение длительного времени. При необходимости работа может выполняться часами.







