


Полная версия
Спортивная психофизиология

Елена Фомина
Спортивная психофизиология: Учебное пособие
Министерство образования и науки Российской Федерации
Федеральное государственное бюджетное образовательное учреждение высшего образования
«Московский педагогический государственный университет»

Рецензенты:
В. Д. Сонькин, доктор биологических наук, профессор, заведующий кафедрой физиологии ФГБОУ ВО РГУФКСМиТ (ГЦОЛИФК)
В. С. Соколова, доктор медицинских наук, профессор кафедры теоретических основ физической культуры и спорта МПГУ
Список сокращений
АТФ – аденозинтрифосфорная кислота
ДНК – дезоксирибонуклеи́новая кислота́
NMDA – рецепторы – рецепторы, активирующиеся
N-метил-D-аспартатом
не-NMDA – рецепторы – рецепторы, не активирующиеся N-метил-D-аспартатом
цАМФ – циклический аденозинмонофосфат
5-НТ – рецептор серотонина
ПКС – протеинкиназа С
ВНД – высшая нервная деятельность
ЧСС – частота сердечных сокращений
ТЛБ – транслокационный белок
БР – белок-регулятор
БМ – белок-м одулятор
РНК – рибонуклеиновая кислота
иРНК – информационная рибонуклеиновая кислота
ДНК – дезоксирибонуклеиновая кислота
ЦНС – центральная нервная система
АКТГ – адренокортикотропный гормон
ЭЭГ – электроэнцефалограмма
G-белки – белки, связывающие гуанозиновые нуклеотиды
Глава 1
Психофизиология памяти и научения. Формирование двигательных навыков
Основные категории обучения
Генетически детерминированные, врожденные формы поведения недостаточны для обеспечения активного существования особи в изменчивой среде. Чем больше изменчивость среды, тем более необходим индивидуальный опыт, в основе которого лежит способность к обучению и память как свойство фиксировать следы обучения.
Обучение – это процесс появления адаптивных изменений поведения в результате приобретенного опыта.
Рассмотрим основные категории обучения.
Неассоциативное, стимулзависимое обучение наблюдается на ранних стадиях онтогенеза, является реакцией на стабильный набор внешних стимулов и не требует ассоциации поведения с внешними сигналами.
Ассоциативное, эффектзависимое обучение возникает на более поздних этапах онтогенеза, когда поведение приобретает активный характер, спектр внешних сигнальных воздействий, ассоциированных с биологическими реакциями, расширяется. Поведение определяется результатом контакта со средой, то есть эффектом взаимодействия со средой.
Когнитивное обучение свойственно взрослым животным с высокоразвитой нервной системой и основано на первых двух категориях обучения. Когнитивное обучение – это свойство формировать функциональную структуру окружающей среды, выявлять закономерности взаимоотношений между компонентами среды.
Неассоциативное обучение
Основными формами неассоциативного, стимулзависимого обучения являются сенситизация, привыкание, импринтинг и подражание.
Сенситизация основана на повышении чувствительности нервной ткани к раздражающим агентам и фасилитации – облегчении запуска реакции, что проявляется в длительном усилении ранее существовавшего ответа животного на раздражитель в результате нанесения болевого стимула.
Время сохранения сенситизации – от нескольких секунд до нескольких недель. Длительные изменения синаптической проводимости, лежащие в основе научения, основаны на тех же механизмах передачи сигнала от нейрона к нейрону. Изучение клеточных основ памяти на млекопитающих представляется довольно затруднительным, в то же время молекулярные механизмы памяти и обучения сходны у простейших животных и у человека. Удобной моделью для изучения механизмов научения является морской моллюск – аплизия.
Сенситизацию можно обнаружить на рефлексе втягивания жабры у аплизии. Биологическое значение сенситизации в том, что животное обучается обращать внимание на стимул, сопровождающийся болью. Когда наносится болевой стимул, рефлекс втягивания жабры на многократную стимуляцию сифона значительно усиливается. Сенситизация влечет за собой изменение синаптической передачи в синапсах, образованных сенсорными нейронами на мотонейронах. Исследования Э. Кендела показали, что нейроны, осуществляющие сенситизацию, усиливают выход медиатора, увеличивая число квантов, высвобождаемых сенсорным нейроном. Другое название этого процесса – пресинаптическое облегчение. Необходимо обратить внимание на то, что на синаптических окончаниях существуют рецепторы, влияющие на высвобождение медиатора. Медиатор модулирующего нейрона – серотонин изменяет, то есть модулирует выделение медиатора сенсорным нейроном.
Э. Кендел предположил, что в головном мозгу имеются синаптические пути, детерминированные процессами развития, которые предрасположены к обучению и могут быть функционально инактивированы или реактивированы опытом. Модификация синапсов происходит под влиянием тренировки или приобретенного опыта. Серотонин связывается с рецептором на пресинаптическом окончании, увеличивая количество С-белка, что в свою очередь увеличивает активность аденилатциклазы. Аденилатциклаза превращает АТФ в цАМФ. Следовательно, под влиянием серотонина в пресинаптическом окончании увеличивается количество цАМФ. Э. Кендел с соавторами указывают на то, что сенситизация сопровождается повышением содержания цАМФ, как и инкубация ганглия с серотонином. В дальнейшем было показано, что введение цАМФ в тело сенсорного нейрона вызывает пресинаптическое облегчение.
Увеличение количества цАМФ ведет к усиленной активации кальциевого тока в результате активации кальциевых каналов или снижению тока калия в результате инактивации калиевых каналов. Приток кальция и выход медиатора повышается с каждым потенциалом действия. Кальций усиливает выделение медиатора, что ведет к проявлению сенситизации.
Механизм действия цАМФ на кальциевые каналы реализуется через протеинкиназы, которые способны фосфорилировать белки, меняя их функцию из-за изменения заряда и конфигурации. Протеинкиназы могут как активировать, так и инактивировать белки канала.
Рассмотрим молекулярные события, приводящие к повышению чувствительности жабры и облегчению синаптического проведения между сенсорным и моторным нейроном (рис. 1). Под буквой А на этом рисунке приведена схема сенситизации жабры в результате приложения повреждающего стимула к хвосту. Стимуляция хвоста активирует сенсорные нейроны, возбуждающие, в свою очередь, фасилитирующие интернейроны. Ряд серотонинергических фасилитирующих нейронов формируют синапсы на терминалях сенсорных нейронов, иннервирующих кожу сифона. Процесс пресинаптической фасилитации ведет к усилению выброса медиатора из сенсорных нейронов.
Схема под буквой Б на рис. 1 отражает биохимические реакции, обеспечивающие пресинаптическую фасилитацию. Действие серотонина на рецептор (5-НТ) приводит к увеличению выброса медиатора.

Рис. 1. Молекулярные события, приводящие к повышению чувствительности жабры (сенситизации) и облегчению (фасилитации) синаптического проведения между сенсорным нейроном и мотонейроном. А – сенситизация жабры приложением повреждающего стимула к другой части тела;Б – пресинаптическая фасилитация в сенсорных нейронах. Пояснения в тексте
Подробнее путь инактивации калиевого канала представлен на рис. 1, он обозначен стрелкой под номером 1. Серотонин связывается с рецептором и увеличивает, как следствие, количество в-белка (Сs), что ведет к повышению активности аденилатциклазы. Аденилатциклаза превращает АТФ в цАМФ и количество цАМФ увеличивается, цАМФ присоединяется к цАМФ-зависимой протеинкиназе и активирует ее каталитическую субъединицу. Каталитическая субъединица, в свою очередь, фосфорилирует калиевый канал, что ведет к изменению его конформации и уменьшению тока через этот канал. Инактивация калиевых каналов затягивает продолжительность потенциала действия, и, как следствие, увеличивается входящий ток ионов кальция, выброс медиатора увеличивается.
Серотонин способен активировать и другой рецептор, увеличивающий количество Сo белка, активирующего фосфолипазу. Этот путь обозначен на рис. 1Б пунктиром. Фосфолипаза стимулирует диацилглицерол в мембране, который активирует протеинкиназу С (ПКС).
Стрелками под номерами 2 и 2а обозначены пути воздействия серотонина, которые опосредованы совместной активностью цАМФ-зависимой протеинкиназы и протеинкиназы С. Эти пути воздействия ведут к увеличению доступности медиатора, благодаря мобилизации синаптических пузырьков из общего пула и усиления экзоцитоза медиатора.
Стрелками под номерами 3 и За обозначены пути усиления выброса медиатора благодаря запуску серотонином цепочек биохимических преобразований, вследствие которых цАМФ-зависимая протеинкиназа и протеинкиназа С открывают кальциевые каналы L-типа.
Общее у сенситизации и условного рефлекса то, что реакция на стимул усиливается в результате активации другого пути. Отличие сенситизации в том, что она не ассоциативна; сенситизирующий стимул повышает реактивность независимо от того, сочетается он со стимулом, вызывающим рефлекс, или нет.
Воздействие повреждающего стимула изучалось с применением болевых воздействий, что ведет к втягиванию жабры. В системе сенсорный нейрон – моторный нейрон, отвечающей за рефлекторное втягивание жабры, важную роль играет модулирующии нейрон, имеющий дополнительный вход к клеточному аппарату, обеспечивающему высвобождение медиатора.
Ведущую роль в обучении играют длительные изменения синаптической проводимости с участием модулирующего нейрона. Формирование связи между двумя нейронами не требует активности постсинаптической клетки, если пресинаптическая клетка модулируется еще одним нейроном, способствующим увеличению выделения медиатора из терминалей пресинаптического нейрона.
Привыкание (габитуация) – устойчивое ослабление реакции в результате многократного действия раздражителя, без сочетания с биологически значимым агентом. Привыкание не является утомлением и зависит от специфичности стимула, силы раздражения и состояния организма.
Многократные предъявления стимула, не имеющего биологического значения, ведут к прекращению ориентировочной реакции. Биологический смысл габитуации – в обеспечении возможности реакции животного на новые стимулы, выделяя такие стимулы на фоне тех, к которым животное уже успело привыкнуть. Отсутствие реакции на знакомые стимулы позволяет экономить энергию. По мере привыкания генерализованная реакция уступает место локальной десинхронизации, длительно сохраняющейся в проекционной зоне анализатора, соответствующего модальности стимула. Привыкание противоположно сенситизации.
Привыкание – самая распространенная форма обучения. Человек, проживающий около железной дороги, перестает реагировать на звук проходящего поезда и во время бодрствования, и во сне. Предъявление иного звука той же интенсивности вызовет пробуждение ото сна, это указывает на специфичность привыкания. Привыкание необходимо отличать от адаптации рецепторов анализатора, когда снижение чувствительности происходит в результате непрерывной стимуляции. Привыкание является особым процессом в нервной системе со своим специфическим механизмом.
Привыкание нейрона – это постепенное ослабление его реакции на повторяющийся раздражитель. Реакция восстанавливается в результате изменения стимула, что указывает на различие с механизмами утомления. Э. Кендел исследовал привыкание на оборонительной реакции аплизии. Слабые тактильные раздражения сифона вызывают рефлекс втягивания жабры. Многократное применение стимуляции сифона моллюска струей холодной воды уменьшает рефлекторное втягивание жабры, наконец втягивание жабры прекращается – произошло привыкание. Повторяющееся возбуждение сенсорных клеток, реагирующих на раздражение сифона моллюска, ведет к инактивации кальциевых каналов и снижению притока ионов Са2 в клетку, как следствие, уменьшается количество медиатора, высвобождаемого в ответ на такое раздражение.
Сенситизация и привыкание являются независимыми процессами и обеспечиваются различными нейронными механизмами. В то же время многократное предъявление раздражителя ведет к сенситизации с последующим подавлением реакции, привыканием, то есть между сенситизацией и привыканием существует тесная взаимосвязь.
Импринтинг (от англ, imprint – отпечатывать, запечатлевать) – комплекс поведенческих реакций новорожденного, обеспечивающих связь между ним и родителями благодаря формированию устойчивой индивидуальной избирательности к внешним стимулам.
Импринтинг замыкает цепь преобразований эмбрионального периода, позволяя новорожденному сформировать механизмы восприятия и реагирования. Основы научной концепции импринтинга заложил в 30-х гг. XX в. Конрад Лоренц. Спустя несколько часов после вылупления утят в инкубаторе К. Лоренц предъявлял им самые разные объекты, например это был сам К. Лоренц, спичечные коробки, ботинки, и птенцы неуклонно следовали за ними, как за матерью-уткой. К. Лоренц считал, что птенцы узнают взрослых представителей своего вида благодаря способности запечатлевать первый движущийся объект, появляющийся в поле зрения после вылупления из яйца, и следуют за ним.
В импринтинге запечатлеваются не индивидуальные, а видоспецифические характеристики объекта. У животных есть определенные генетические предпосылки для научения поведенческим актам, свойственным своему виду. Если птенец белоголовой воробьиной овсянки будет слышать только американского певчего воробья, то, став взрослым, он не сможет петь песню своего вида. Генетические предпосылки для песни американского певчего воробья у овсянки отсутствуют, в связи с этим ее песня не будет в точности такой, как у певчего воробья.
Импринтинг происходит только в период созревания сенсорных систем организма. К. Лоренц выделил ряд особенностей, отличающих импринтинг от ассоциативного поведения:
– наличие критического периода (ограниченного временного отрезка), чаще всего он наступает после рождения, но сроки критического периода могут быть и иными, это определяется степенью созревания соответствующих сенсорных систем;
– сохранение на всю жизнь без подкрепления;
– отсутствие подкрепления в виде биологически значимого стимула.
Детальное изучение импринтинга (реакции следования) проводилось с помощью специальной установки, где утенок следовал за моделью утки (рис. 2). Оказалось, что время экспозиции объекта не имеет значения. Явление импринтинга оказалось значительно шире, чем его рассматривал К. Лоренц. К импринтингу относят также устойчивое запоминание животным места своего рождения (хоминг), карты звездного неба и других жизненно важных ориентиров. Считают, что моторным эквивалентом реакции следования у человека является улыбка ребенка и появление комплекса оживления на втором месяце жизни, то есть интенсивные двигательные реакции при узнавании лиц близких людей.
Если зрительные объекты запечатлеваются после рождения, то звуковые сигналы могут импринтироваться и до рождения. Успокаивающее действие на новорожденного оказывает воспроизведение звукозаписи сердцебиения матери в спокойном состоянии. Рекомендация беременным женщинам разговаривать с будущим ребенком не только на родном языке основана на явлении импринтинга.
Интересно также явление «полового импринтирования», описаное К. Лоренцом. Самец птицы, воспитанной особями другого вида, предпочитает в качестве полового партнера самку, принадлежащую виду приемных родителей. Птенцы, участвовавшие в экспериментах К. Лоренца, искали брачных партеров, сходных с человеком. Кот, воспитанный собакой, направляет свое половое поведение на собаку.

Рис. 2. Прибор, применяемый для изучения импринтинга
В последнее время внимание исследователей поведения привлекают феромоны – биологически активные вещества, синтезируемые специальными органами и выделяемые во внешнюю среду. Возможно, механизм влияния феромонов на взаимоотношения детей и родителей также основан на импринтинге. Феромоны матери означают для младенца спокойствие и безопасность. Современная акушерская практика рекомендует контакт матери и новорожденного сразу после родов. В Древнем Риме отец должен был взять новорожденного на руки, а на Руси новорожденного заворачивали в рубаху, только что снятую с отца. Эти действия позволяют новорожденному навсегда запечатлеть феромоны родителей.
Подражание – выполнение действий в результате наблюдения за животными своего вида. Имитационное поведение имеет охранительное значение. Гибель или повреждение одного из членов сообщества ведет к формированию у свидетелей события защитного поведения, позволяющего избежать опасности. Подражание побуждает к одновременному собиранию пищи, спасению от хищника, отдыху, игровому поведению детенышей.
К подражанию можно отнести и обучение путем наблюдения, в котором выделяют два вида: простое наблюдение и викарное научение. Когда обезьяна обучается мыть банан, наблюдая за исследователем, без соответствующих подкреплений, это можно отнести к простому подражанию, так как обезьяна не понимает, зачем она это делает. Викарное научение – это наблюдение и подражание, когда важен результат действия. В раннем онтогенезе человек обычно использует обучение путем наблюдения, с возрастом начинает превалировать викарное обучение.
Ассоциативное обучение
Ассоциативное обучение отличается от неассоциативного наличием подкрепления, в роли которого может выступить безусловное раздражение. Первоначально рассмотрим клеточные аналоги ассоциативного обучения. Ассоциация нейронов обеспечивается конвергенцией подкрепляющего раздражителя и условного сигнала. Этот процесс назван сейчас премодулирующим ассоциативным механизмом. Э. Кэндэл описал клеточный уровень выработки условного рефлекса на базе безусловного рефлекса втягивания жабры морской улиткой аплизией. Обычно рефлекторное втягивание жабры происходит при раздражении тела с противоположной от жабры стороны, что ведет к стимуляции сенсорных нейронов, связанных с сифоном или с мантийным выступом (рис. 3). Сенсорные нейроны контактируют с мотонейронами жабры и интернейронами, образующими синапсы на этих же мотонейронах. Рефлекс втягивания жабры может быть вызван и раздражением хвоста.
Раздражение хвоста аплизии электрическим током было выбрано в качестве безусловного раздражителя при выработке условного рефлекса. Слабое раздражение нервного пути, обслуживающего сифон, было выбрано в качестве условного раздражителя. Слабое тактильное раздражение сифона сочетали с ударом электрическим током по хвостовой части аплизии. В качестве контроля использовали нервный путь, обслуживающий мантию. Контрольный нервный путь раздражали столько же раз, но без электрического удара. После пяти сочетаний условного и безусловного стимула ответ на раздражение сифона становится сильнее, чем ответ на раздражение мантии, то есть вырабатывается дифференцировка, запоминается подкрепляемое поведение. После выработки условного рефлекса даже слабая стимуляция сенсорных нейронов сифона вызывает возбуждающие постсинаптические потенциалы в мотонейронах и интернейронах и ведет к условно-рефлекторному втягиванию жабры.
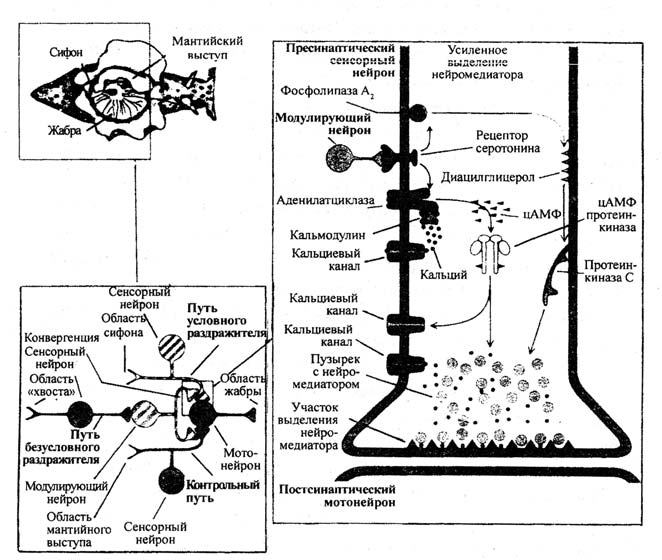
Рис. 3. Условные рефлексы у аплизии. Нервный путь, участвующий в условном рефлексе втягивания жабры у Ар1узт (внизу слева). Справа молекулярные механизмы повышения проводимости через синапс
Безусловное подкрепляющее раздражение «хвоста» активирует многие группы клеток, включая те, которые отвечают за движение жабры, вызывая усиление выделения нейромедиатора из терминалей сенсорных нейронов. Такое усиление выброса медиатора обеспечивается влиянием модулирующих нейронов.
Механизм сенситизации, обусловленный усилением выброса медиатора под влиянием модулирующего нейрона, был рассмотрен нами выше. В случае ассоциативного обучения сенсорный нейрон возбуждается под действием условного раздражителя на сифон наряду с воздействием безусловного раздражителя через модулирующие нейроны. В данном случае условный и безусловный раздражители представлены внутри одной клетки конвергенцией двух сигналов на одном и том же ферменте – аденилатциклазе. Влияние модулирующего нейрона через серотонинергический механизм сочетается с влиянием условного раздражителя через сенсорный нейрон.
Стимуляция одних модулирующих нейронов вызывает меньшее синаптическое облегчение, чем после воздействия на сенсорный пресинаптический нейрон условного раздражителя.
Классические условные рефлексы относятся к ассоциативному обучению, то есть обучение характеризуется ассоциацией во времени индифферентного раздражителя с деятельностью организма.
Структурно-функциональной основой условного рефлекса служат кора и подкорковые образования мозга. Условно-рефлекторная деятельность сводится к превращению индифферентного раздражителя в сигнальный благодаря многократному подкреплению раздражения безусловным стимулом. Эффекторным звеном рефлекторной дуги условного рефлекса может выступать любой иннервируемый орган.
Образование условного рефлекса происходит с соблюдением нескольких правил:
• индифферентный раздражитель должен иметь достаточную силу для возбуждения определенных рецепторов;
• индифферентный раздражитель должен предшествовать безусловному либо предъявляться одновременно с ним;
• раздражитель, используемый в качестве условного, должен быть слабее безусловного;
• необходимо нормальное физиологическое состояние корковых и подкорковых структур, образующих центральное представительство соответствующего условного и безусловного стимулов;
• отсутствие сильных посторонних раздражителей;
• отсутствие значительных патологических процессов в организме.
И. П. Павлов первоначально предполагал, что условный рефлекс образуется на уровне кора – подкорковые образования (временная связь замыкается между корковыми нейронами в зоне представительства индифферентного условного стимула и подкорковыми нервными клетками, составляющими центральное представительство безусловного раздражителя). Позднее И. П. Павлов образование условно-рефлекторной связи объяснял образованием связи на уровне корковых зон представительства условного и безусловного стимулов.
Современные данные указывают на возможность разных уровней замыкания, формирования условно-рефлекторной связи (кора – кора, кора – подкорковые образования, подкорковые образования – подкорковые образования) при доминирующей роли в этом процессе корковых структур. Очевидно, физиологический механизм образования условного рефлекса представляет собой сложную динамическую организацию корковых и подкорковых структур мозга.
Классические исследования условно-рефлекторной деятельности организма были проведены И. П. Павловым с использованием рефлексов слюноотделения (эффекторное звено рефлекторной дуги – слюнные железы). Животному предъявляют индифферентный по отношению к безусловному рефлексу слюноотделения стимул с последующим или одновременным подкреплением его безусловным раздражителем. Предварительно производят операцию выведения протока слюнной железы на наружную поверхность кожи.
Впоследствии стали широко применять и методики изучения двигательных условных рефлексов с эффекторным звеном рефлекторной дуги, образованным мышцами, обеспечивающими двигательные акты.
Стадии формирования условного рефлекса
В формировании условного рефлекса различают две стадии: начальную стадию генерализации условного возбуждения и конечную стадию концентрации условного возбуждения.
Начальная стадия генерализованного условного возбуждения является продолжением более общей универсальной реакции организма на любой новый для него раздражитель, представленной ориентировочным рефлексом.
Эта стадия образования условного рефлекса состоит в формировании временной связи не только на данный конкретный условный раздражитель, но и на все близкие ему стимулы. Нейрофизиологический механизм заключается в иррадиации возбуждения из центра проекции условного раздражителя на нервные клетки окружающих проекционных зон, близких в функциональном отношении клеткам центрального представительства условного раздражителя. Условно-рефлекторный ответ наблюдается на сходные, близкие стимулы как результат распространения возбуждения из проекционной зоны основного условного стимула, например реакция слюноотделения на звук, близкий к условному по тону.
