


Полная версия
Физиология человека. Общая. Спортивная. Возрастная: учебник, 7-е издание
Развивая идеи И. М. Сеченова о рефлекторной основе поведенческой деятельности целостного организма, И. П. Павлов пришел к мысли, что в изменяющихся условиях внешней среды недостаточно обладать стандартными рефлекторными реакциями, а требуется выработка новых рефлексов, адекватных новым условиям существования. Впервые об условных рефлексах он заговорил в известной Мадридской речи в 1903 г.
4.1. Условия образования и разновидности условных рефлексов
Условные рефлексы по многим признакам отличаются от безусловных (табл. 1).
Условные рефлексы у млекопитающих и человека осуществляются корой больших полушарий (в этом также принимают участие таламический отдел промежуточного мозга и в ряде случаев подкорковые ядра).
И. П. Павловым была разработана объективная методика изучения приобретаемых или условных рефлексов, которая основывалась на изоляции обследуемого организма от посторонних раздражений и на точной регистрации сигнала и ответа на него. Исследования проводились на собаках в звукоизолированных камерах («башнях молчания»), куда подавались дозированные раздражения светом, звуком, механическими раздражениями кожи и пр. В качестве ответной реакции было выбрано выделение слюны, которая отводилась от одного из слюнных протоков, выведенного на наружную поверхность щеки (методика фистулы слюнной железы).
Таблица 1
Различия условных и безусловных рефлексов

В процессе выработки приобретаемых рефлексов должны соблюдаться следующие условия:
• сочетание любого индифферентного раздражителя с каким-либо значимым безусловным раздражением (например, пищевым) – методика безусловного подкрепления;
• индифферентное раздражение должно предшествовать безусловному, чтобы приобрести сигнальное значение;
• нервные центры, к которым адресованы раздражения, должны быть в состоянии оптимального возбуждения.
Например, после предварительного изолированного действия светового сигнала собаке подавалось подкрепление – мясосухарный порошок и регистрировалось выделение слюны. После ряда сочетаний этих сигналов уже одно только включение света вызывало выделение слюны, т. е. был выработан новый рефлекс, биологический смысл которого заключался в подготовке организма к приему пищи.
Механизм образования условного рефлекса заключался в формировании новой рефлекторной дуги, в которой к эфферентной части безусловного рефлекса присоединялась новое афферентное начало рефлекторной дуги, идущее от зрительных путей. Между центрами этих исходных рефлексов сформировалась новая связь, которую И. П. Павлов назвал временной связью, так как в случае прекращения подачи пищи после светового сигнала слюнной условный рефлекс исчезал.
В ходе выработки условного рефлекса наблюдались определенные фазы этого процесса:
1) генерализации (обобщенное восприятие сигнала, когда условная реакция наблюдалась на любой сходный сигнал), основой чего были процессы иррадиации возбуждения в коре больших полушарий;
2) концентрации возбуждения (реакция только на конкретный сигнал), что появлялось за счет вырабатываемого условного торможения на посторонние неподкрепляемые сигналы;
3) стабилизации (упрочения условного рефлекса).
В дальнейших исследованиях условные рефлексы были выработаны в разнообразных экспериментальных условиях (в том числе в условиях свободного поведения) у различных животных, птиц, рыб, черепах, даже у амеб. Изучение биопотенциалов коры больших полушарий показало, что условием образования временной связи между изучаемыми корковыми центрами является пространственная синхронизация их электрической активности.
Различают несколько разновидностей условных рефлексов:
1) натуральные – на сигналы, характеризующие безусловные раздражители (например, запах мяса для слюнного рефлекса), и искусственные – на посторонние сигналы (например, запах мяты);
2) наличные и следовые на условный сигнал, непосредственно предшествующий безусловному подкреплению, и на его следовое влияние;
3) положительные (с активным проявлением ответной реакции) и отрицательные (с ее торможением);
4) условные рефлексы на время – при ритмической подаче условных сигналов ответная реакция появляется через заданный интервал даже при отсутствии очередного сигнала;
5) условные рефлексы первого порядка – на один предшествующий условный раздражитель – и более высоких порядков, когда безусловному подкреплению предшествует сочетание двух последовательно подающихся сигналов (свет + звук) – условный рефлекс второго порядка, трех сигналов (свет + звук + касалка) – условный рефлекс третьего порядка и т. д.
У собак вырабатываются в основном рефлексы третьего порядка, у обезьян – четвертого, у грудного ребенка – 5–6 порядка, у взрослого человека – двадцатого и более порядков. Освоение речи человеком представляет собой формирование огромной цепи условно-безусловных рефлексов, не требующих специального подкрепления.
При формировании новых двигательных навыков возникают особые рефлексы, которые в отличие от сенсорных рефлексов или рефлексов I рода (в которых новой частью рефлекторной дуги была афферентная часть) имеют новую часть рефлекторной дуги в эфферентном отделе (новые исполнительные аппараты – мышцы). Это так называемые инструментальные, или оперантные, рефлексы – рефлексы II рода (Конорский Ю. М., 1970).
4.2. Внешнее и внутреннее торможение условных рефлексов
По своему происхождению торможение условных рефлексов может быть безусловным (врожденным) и условным (выработанным в течение жизни). К безусловному торможению относят охранительное, или запредельное, торможение, возникающее при чрезмерно сильном или длительном раздражении, и внешнее торможение условных рефлексов посторонними для центров условного рефлекса раздражителями (например, нарушение непрочного двигательного навыка у спортсмена в необычных условиях соревнований).
Условное торможение вырабатывается при отсутствии подкрепления условного сигнала. Различают несколько видов условного торможения: угасательное, дифференцировочное и запаздывающее.
Угасание развивается при повторении условного сигнала без подкрепления. Например, имея прочный слюнный условный рефлекс у собаки на вспышку света и затем применяя свет без подкрепления, можно получить последовательно следующие условные ответы – 10, 8, 6, 4, 5, 2, 0, 0, 0 капель слюны.
Дифференцировочное торможение вырабатывается при подкреплении одного условного сигнала (например, звук с частотой 500 Гц) и отсутствии подкрепления сходных с ним сигналов (звук 1000, 200 и 100 Гц), на которые первоначально (в период генерализации условного рефлекса) получался условный ответ. Этот вид торможения, в частности, позволяет спортсмену отдифференцировать сокращения ненужных мышц при выработке двигательного навыка, т. е. имеет важное координационное значение. Процесс воспитания человека сопровождается постоянной дифференцировкой подкрепляемых и осуждаемых обществом поведенческих реакций (что такое «хорошо» и что такое «плохо»).
Запаздывающее торможение формируется при отставлении на определенный отрезок времени подкрепления от условного сигнала. В этом случае сразу после условного сигнала реакция отсутствует (тормозится), но перед моментом подкрепления обнаруживается.
4.3. Динамический стереотип
В жизни обычно встречаются не отдельные условные рефлексы, а сложные их комплексы, в которых они сочетаются с безусловными рефлексами (двигательными, сердечно-сосудистыми, дыхательными и пр.). Систему условных и безусловных рефлексов И. П. Павлов назвал динамическим стереотипом. Она вырабатывается при повторении одного и того же порядка раздражений (ситуаций) и соответственно выражается в цепи закрепленных ответных реакций, т. е. стереотипе. Но изменение внешних условий может вызвать перестройку этой системы или ее разрушение, что отмечается термином «динамический».
Например, у собаки выработан динамический стереотип на определенный порядок из 6 раздражителей, и имеются на них закрепленные условные величины слюноотделения, специфические для каждого сигнала: 1) свет – 12 капель; 2) звук – 20 капель; 3) метроном 120 уд./с – 10 капель; 4) метроном 60 уд./с (неподкрепляемый раздражитель) – 0 капель; 5) свет – 12 капель; 6) звук – 20 капель. Если теперь подавать один и тот же сигнал, то ответная цепь реакций сохранится прежней: 1) свет – 12 капель; 2) свет – 20 капель; 3) свет – 10 капель; 4) свет – 0 капель; 5) свет – 12 капель; 6) свет – 20 капель. Однако изолированное включение светового раздражения сохраняет обычный ответ – 12 капель.
Следовательно, в коре больших полушарий собаки образована цепь последовательно возбуждающихся или затормаживающихся нервных центров, в которой активность каждого автоматически вызывает включение следующего. Подобный стереотип возникает у спортсмена при выработке двигательного навыка, особенно при выполнении стандартных движений. Такой стереотип, связанный с цепью моторных актов, А. Н. Крестовников назвал «двигательный динамический стереотип». Он легче образуется при выполнении циклических упражнений, чем ациклических.
4.4. Типы высшей нервной деятельности, первая и вторая сигнальная система
Случившееся в 1924 г. в Ленинграде сильное наводнение грозило затопить клетки с подопытными собаками, которые испытали сильный стресс. На следующий день обнаружилось, что у некоторых из них пропали прочно выработанные условные рефлексы, но у других рефлексы сохранились. Это навело И. П. Павлова на мысль о различных типах нервной системы у животных. В качестве основных свойств нервной системы И. П. Павлов рассматривал силу возбуждения и торможения, их уравновешенность и подвижность. С учетом этих свойств им были выделены следующие четыре типа высшей нервной деятельности (ВНД), которые оказались сходными с четырьмя темпераментами, выделенными еще Гиппократом в V веке до н. э.
1. Тип сильный неуравновешенный (холерик). Характеризуется сильным процессом возбуждения и более слабым процессом торможения, поэтому легко возбуждается и с трудом затормаживает свои реакции.
2. Тип сильный уравновешенный и высокоподвижный (сангвиник). Отличается сильными уравновешенными и высокоподвижными процессами возбуждения и торможения. Легко переключается с одной формы деятельности на другую, быстро адаптируется к новой ситуации.
3. Тип сильный уравновешенный инертный (флегматик). Имеет сильные и уравновешенные процессы возбуждения и торможения, но малоподвижный – медленно переключающийся с возбуждения на торможение и обратно. С трудом переходит от одного вида деятельности к другому, зато вынослив при длительной работе. Медленно, но прочно адаптируется к необычным условиям внешней среды.
4. Тип слабый (меланхолик). Характеризуется слабыми процессами возбуждения и торможения, с некоторым преобладанием тормозного процесса, мало адаптивен, подвержен неврозам. Зато обладает высокой чувствительностью к слабым раздражениям и может их легко дифференцировать.
Описанные типы имеются у животных и человека. Они представляют собой лишь крайние проявления особенностей нервной системы, между которыми может быть значительное число переходных типов.
Кроме того, И. П. Павлов выделил специфически человеческие типы ВНД, связанные с наличием у человека особой второй сигнальной системы – слова видимого, слышимого, написанного, произносимого, в отличие от первой сигнальной системы, общей для человека и животных, – непосредственных раздражителей внешней или внутренней среды организма. Вторая сигнальная система чрезвычайно расширила адаптационные возможности человека. Ее свойствами являются: обобщение сигналов первой и второй сигнальной системы, появление абстракций (сложных комплексных понятий – мужество, ярость, доброта и пр.), возможность передачи накопленного опыта предшествующих поколений последующим (возникновение науки, культуры и пр.). Вторая сигнальная система таким образом составила основу письменной и устной речи, появления математических и нотных символов, абстрактного мышления человека. Ее деятельность связывают с функциями третичных полей коры больших полушарий, преимущественно левого полушария у правшей, где находятся центры речи.
В связи с различным соотношением у людей реакций, связанных с преобладанием первой или второй сигнальной системы, И. П. Павлов различал специфически человеческие типы нервной системы: «мыслительный» – с преобладанием второй сигнальной системы; «художественный» – с преобладанием первой сигнальной системы. Среди взрослых людей количество лиц с преобладанием второй сигнальной системы составляет около половины населения. Около 25 % составляют лица с преобладанием первой сигнальной системы и примерно 25 % – лица, имеющие равновесие обеих систем. Соответственно этим типам в настоящее время различают две основные формы интеллекта человека: невербальный интеллект, отражающий природные возможности индивида манипулировать с непосредственными (особенно зрительно-пространственными) раздражителями, и вербальный интеллект, отражающий способность манипулировать со словесным материалом, что определяет характер поведенческих реакций, в том числе и в спорте.
5. Нервно-мышечный аппарат
У человека существует три вида мышц: поперечно-полосатые скелетные мышцы, особая поперечно-полосатая сердечная мышца и гладкие мышцы внутренних органов.
5.1. Функциональная организация скелетных мышц
Скелетные мышцы человека содержат около 300 млн мышечных волокон и имеют площадь порядка 3 м2. Целая мышца представляет собой отдельный орган, а мышечное волокно – клетку. Мышцы иннервируются двигательными нервами, передающими из центров моторные команды, чувствительными нервами, несущими в центры информацию о напряжении и движении мышц, и симпатическими нервными волокнами, влияющими на обменные процессы в мышце. Функции скелетных мышц заключаются в перемещении частей тела друг относительно друга, перемещении тела в пространстве (локомоция) и поддержании позы тела.
Функциональной единицей мышцы является двигательная единица, состоящая из мотонейрона спинного мозга, его аксона (двигательного нерва) с многочисленными окончаниями и иннервируемых им мышечных волокон. Возбуждение мотонейрона вызывает одновременное сокращение всех входящих в эту единицу мышечных волокон. Двигательные единицы (ДЕ) небольших мышц содержат малое число мышечных волокон (ДЕ мышц глазного яблока 3–6 волокон, мышц пальцев руки 10–25 волокон), а ДЕ крупных мышц туловища и конечностей – до нескольких тысяч (например, ДЕ икроножной мышцы человека – около 2000 мышечных волокон).
Мелкие мышцы иннервируются из одного сегмента спинного мозга, а крупные мышцы – мотонейронами 2–3 спинальных сегментов. Мотонейроны, иннервирующие одну мышцу, составляют общий мотонейронный пул, в котором могут находиться мотонейроны различных размеров. Большие ДЕ образованы крупными мотонейронами, которые имеют толстые аксоны, множество концевых разветвлений и большое число связанных с ними мышечных волокон. Такие ДЕ имеют низкую возбудимость, генерируют высокую частоту нервных импульсов (порядка 20–50 импульсов в 1 с) и характеризуются высокой скоростью проведения возбуждения. Они включаются в работу лишь при высоких нагрузках на мышцу. Мелкие ДЕ имеют мотонейроны небольших размеров, тонкие и медленно проводящие аксоны, малое число мышечных волокон. Они легко возбудимы и включаются в работу при незначительных мышечных усилиях. Нарастание нагрузки вызывает активацию различных ДЕ скелетной мышцы в соответствии с их размерами – от меньших к большим (правило Хеннемана).
Мышечное волокно представляет собой вытянутую клетку (ее диаметр около 10–100 мкм, а длина 10–12 см). В состав волокна входят его оболочка – сарколемма, жидкое содержимое – саркоплазма, ядро, энергетические центры – митохондрии, белковые депо – рибосомы, сократительные элементы – миофибриллы, а также замкнутая система продольных трубочек и цистерн, расположенных вдоль миофибрилл и содержащих ионы Са2+, – саркоплазматический ретикулум. Поверхностная мембрана клетки через равные промежутки образует поперечные трубочки, входящие внутрь мышечного волокна, по которым внутрь клетки проникает потенциал действия при ее возбуждении.
Миофибриллы – это тонкие волокна (диаметр 1–2 мкм, длина 2–2,5 мкм), содержащие два вида сократительных белков (протофибрилл): тонкие нити актина и вдвое более толстые нити миозина. Они расположены таким образом, что вокруг миозиновых нитей находится 6 актиновых нитей, а вокруг каждой актиновой – 3 миозиновых. Миофибриллы разделены Z-мембранами на отдельные участки – саркомеры, в средней части которых расположены преимущественно миозиновые нити, а актиновые нити прикреплены к Z-мембранам по бокам саркомера. (Разная способность актина и миозина преломлять свет создает в состоянии покоя мышцы ее поперечно-полосатый вид в световом микроскопе.)
Нити актина составляют около 20 % сухого веса миофибрилл. Актин состоит из двух форм белка: 1) глобулярной формы – в виде сферических молекул и 2) палочковидных молекул тропомиозина, скрученных в виде двунитчатых спиралей в длинную цепь. На протяжении этой двойной актиновой нити каждый виток содержит по 14 молекул глобулярного актина (по 7 молекул с обеих сторон), наподобие нитки с бусинками, а также центры связывания ионов Са2+. В этих центрах содержится особый белок (тропонин), участвующий в образовании связи актина с миозином.
Миозин составлен из уложенных параллельно белковых нитей (эта часть представляет собой так называемый легкий меромиозин). На обоих концах его имеются отходящие в стороны шейки с утолщениями – головками (эта часть – тяжелый меромиозин), благодаря которым образуются поперечные мостики между миозином и актином.
5.2. Механизмы сокращения и расслабления мышечного волокна
При произвольной внутренней команде сокращение мышцы человека начинается примерно через 0,05 с (50 мс). За это время моторная команда передается от коры больших полушарий к мотонейронам спинного мозга и по двигательным волокнам к мышце. Подойдя к мышце, процесс возбуждения должен с помощью медиатора преодолеть нервно-мышечный синапс, что занимает примерно 0,5 мс. Медиатором здесь является ацетилхолин, который содержится в синаптических пузырьках в пресинаптической части синапса. Нервный импульс вызывает перемещение синаптических пузырьков к пресинаптической мембране, их опорожнение и выход медиатора в синаптическую щель. Действие ацетилхолина на постсинаптическую мембрану чрезвычайно кратковременно, после чего он разрушается ацетилхолинэстеразой на уксусную кислоту и холин. По мере расходования запасы ацетилхолина постоянно пополняются путем его синтезирования в пресинаптической мембране. Однако при очень частой и длительной импульсации мотонейрона расход ацетилхолина превышает его пополнение, а также снижается чувствительность постсинаптической мембраны к его действию, в результате чего нарушается проведение возбуждения через нервно-мышечный синапс. Эти процессы лежат в основе периферических механизмов утомления при длительной и тяжелой мышечной работе.
Выделившийся в синаптическую щель медиатор прикрепляется к рецепторам постсинаптической мембраны и вызывает в ней явления деполяризации. Небольшое подпороговое раздражение вызывает лишь местное возбуждение небольшой амплитуды – потенциал концевой пластинки (ПКП).
При достаточной частоте нервных импульсов ПКП достигает порогового значения и на мышечной мембране развивается мышечный потенциал действия. Он (со скоростью 5 м/с) распространяется вдоль по поверхности мышечного волокна и заходит в поперечные трубочки внутрь волокна. Повышая проницаемость клеточных мембран, потенциал действия вызывает выход из цистерн и трубочек саркоплазматического ретикулума ионов Са2+, которые проникают в миофибриллы, к центрам связывания этих ионов на молекулах актина.
Под влиянием Са2+ длинные молекулы тропомиозина проворачиваются вдоль оси и скрываются в желобки между сферическими молекулами актина, открывая участки прикрепления головок миозина к актину. Тем самым между актином и миозином образуются так называемые поперечные мостики. При этом головки миозина совершают гребковые движения, обеспечивая скольжение нитей актина вдоль нитей миозина с обоих концов саркомера к его центру, т. е. механическую реакцию мышечного волокна (рис. 10).
Энергия гребкового движения одного мостика производит перемещение на 1 % длины актиновой нити. Для дальнейшего скольжения сократительных белков друг относительно друга мостики между актином и миозином должны распадаться и вновь образовываться на следующем центре связывания Са2+. Такой процесс происходит в результате активации в этот момент молекул миозина. Миозин приобретает свойства фермента АТФазы, который вызывает распад АТФ. Выделившаяся при распаде АТФ энергия приводит к разрушению имеющихся мостиков и образованию в присутствии Са2+ новых мостиков на следующем участке актиновой нити. В результате повторения подобных процессов многократного образования и распада мостиков сокращается длина отдельных саркомеров и всего мышечного волокна в целом. Максимальная концентрация кальция в миофибрилле достигается уже через 3 мс после появления потенциала действия в поперечных трубочках, а максимальное напряжение мышечного волокна – через 20 мс.
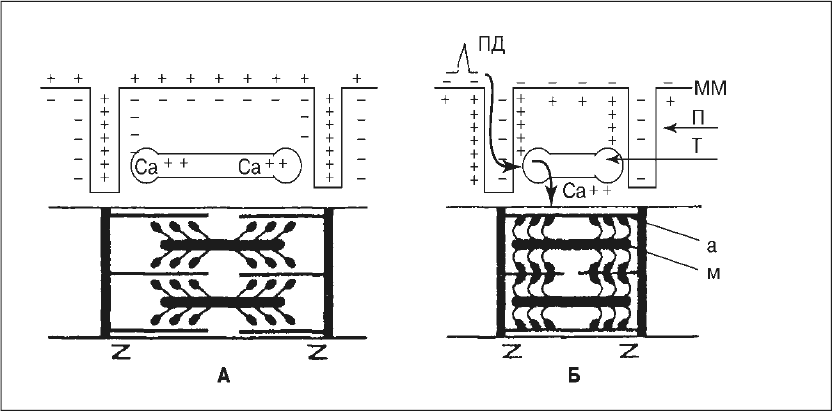
Рис. 10. Схема электромеханической связи в мышечном волокне. А – состояние покоя; Б – возбуждение и сокращение; ПД – потенциал действия; ММ – мембрана мышечного волокна; П – поперечные трубочки; Т – продольные трубочки и цистерны с ионами Са2+; а – тонкие нити актина, м – толстые нити миозина с утолщениями (головками) на концах. Зет-мембранами ограничены саркомеры миофибрилл. Толстые стрелки – распространение потенциала действия при возбуждении волокна и перемещение ионов Са2+ из цистерн и продольных трубочек в миофибриллы, где они содействуют образованию мостиков между нитями актина и миозина и скольжение этих нитей (сокращение волокна) за счет гребковых движений головок миозина
Весь процесс от появления мышечного потенциала действия до сокращения мышечного волокна называется электромеханической связью (или электромеханическим сопряжением). В результате сокращения мышечного волокна актин и миозин более равномерно распределяются внутри саркомера, и исчезает видимая под микроскопом поперечная исчерченность мышцы.
Расслабление мышечного волокна связано с работой особого механизма – «кальциевого насоса», который обеспечивает откачку ионов Са2+ из миофибрилл обратно в трубочки саркоплазматического ретикулума. На это также тратится энергия АТФ.
5.3. Одиночное и тетаническое сокращение. Электромиограмма
При единичном надпороговом раздражении двигательного нерва или самой мышцы возбуждение мышечного волокна сопровождается одиночным сокращением. Эта форма механической реакции состоит из трех фаз: латентного, или скрытого, периода; фазы сокращения; фазы расслабления. Самой короткой фазой является скрытый период, когда в мышце происходит электромеханическая передача. Фаза расслабления обычно в 1,5–2 раза более продолжительна, чем фаза сокращения, а при утомлении затягивается на значительное время.
Если интервалы между нервными импульсами короче, чем длительность одиночного сокращения, то возникает явление суперпозиции – наложение механических эффектов мышечного волокна друг на друга и наблюдается сложная форма сокращения – тетанус. Различают две формы тетануса – зубчатый тетанус, возникающий при более редком раздражении, когда происходит попадание каждого следующего нервного импульса в фазу расслабления отдельных одиночных сокращений, и сплошной, или гладкий, тетанус, возникающий при более частом раздражении, когда каждый следующий импульс попадает в фазу сокращения (рис. 11). Таким образом (в некоторых границах) между частотой импульсов возбуждения и амплитудой сокращения волокон ДЕ существует определенное соотношение: при небольшой частоте (например, 5–8 имп. в 1 с) возникают одиночные сокращения, при увеличении частоты (15–20 имп. в 1 с) – зубчатый тетанус, при дальнейшем нарастании частоты (25–60 имп. в 1 с) – гладкий тетанус. Одиночное сокращение – более слабое и менее утомительное, чем тетаническое. Зато тетанус обеспечивает в несколько раз более мощное, хотя и кратковременное сокращение мышечного волокна.


