


Полная версия
Болезни суставов
В эпифизах окостенение начинается позже, у некоторых костей даже после рождения. Окостенение начинается из костной точки, появляющейся внутри хрящевой закладки эпифиза. Данный процесс окостенения называют энхондральным. Он проходит следующим образом. Вначале из надкостницы вглубь хряща по радиусам прорастают кровеносные сосуды. В самой середине эпифиза хрящ обызвествляется и рассасывается, а на его месте развивается костная ткань. Позже за счет надкостницы по краю хрящевой закладки эпифиза развивается периостальная кость (перихондральная), которая представлена тонкой пластинкой компактного вещества. Перихондральная пластинка отсутствует лишь в области будущих суставных поверхностей костей, там остается хорошо выраженный слой хряща. Хрящевая прослойка остается также между эпифизом и диафизом, это метаэпифизарный хрящ. Он является зоной роста кости в длину и исчезает лишь после прекращения роста кости.
У длинных трубчатых костей (бедро, кости голени, плечевая кость, кости предплечья) обычно формируются отдельные точки окостенения в каждом эпифизе. Прирастание эпифизов к диафизу обычно происходит после рождения. Так, у большеберцовой кости нижний эпифиз прирастает к 22 годам, а верхний – к24годам. У коротких трубчатых костей (кости пясти, плюсны, фаланги), как правило, имеется точка окостенения только в одном эпифизе, а другой эпифиз окостеневает за счет диафиза. У некоторых трубчатых костей в эпифизе появляется одновременно несколько точек окостенения, например, в верхнем эпифизе плеча – три точки, в нижнем – четыре.
Кости с объемной формой (кости запястья, предплюсны) окостеневают так же, как и эпифизы длинных трубчатых костей, т. е. энхондральное окостенение предшествует периостальному. В плоских костях процесс идет противоположно, т. е. периостальное окостенение предшествует энхондральному.
Следует обратить внимание на то, что кроме главных точек окостенения могут быть добавочные. Они появляются значительно позже главных. С наступлением полового созревания метаэпифизарные хрящи истончаются и замещаются костной тканью. В скелете образуются синостозы. Первыми прирастают дистальный эпифиз плечевой кости и эпифизы пястных костей. Завершается образование синостозов к 24–25 годам. Рост кости заканчивается в тот момент, когда все главные и добавочные точки сливаются в одну массу, т. е. после исчезновения хрящевых прослоек, отделяющих части кости друг от друга.
Наблюдаются значительные индивидуальные различия в темпах окостенения. Процесс окостенения скелета у ребенка может ускоряться или замедляться, что обусловлено генетическими, гормональными и средовыми факторами. Для оценки процесса развития скелета у ребенка введено понятие «костный возраст», о котором судят по числу имеющихся в костях точек окостенения и срокам их слияния. Для определения окостенения обычно производят рентгеновские снимки кисти, так как в этой части тела особенно четко выявляется возрастная динамика появления точек окостенения и развития синостозов. Так, для костей запястья характерны следующие сроки появления точек окостенения: у новорожденного все запястье хрящевое; на 1-м году образуются точки окостенения в головчатой и крючковидной кости; на 3-м – в трехгранной; на 4-м – вполулунной; на 5-м – в ладьевидной; на 6–7-м – в кости трапеции и в трапециевидной кости; на 10–14-м – в гороховидной кости.
В. С. Сперанский (1998) выделяет следующие закономерности процесса окостенения:
1) в перепончатой основе (соединительнотканной) окостенение начинается раньше, чем в хряще;
2) окостенение скелета происходит в краниокаудальном направлении;
3) в черепе окостенение распространяется от лицевого черепа к мозговому;
4) в свободных конечностях окостенение идет от проксимальных отделов к дистальным.
Костный возраст не всегда совпадает с паспортным. Так, у одних детей процесс окостенения завершается на 1–2 года раньше положенного срока, у других – на 1–2 года отстает.
Начиная с 9 лет, отчетливо выявляются половые различия окостенения: у девочек этот процесс происходит быстрее. Рост тела в длину у девушек завершается в 16–17 лет, у юношей – в 17–18 лет. После этого возраста прирост длины тела составляет не более 2 %.
При старении в различных частях скелета происходит разрежение кости – остеопороз. В трубчатых костях отмечается рассасывание кости на внутренней поверхности диафиза, в результате чего расширяется костномозговая полость.
Вместе с этим наблюдается обызвествление и развитие костной ткани на внешней поверхности костей, под надкостницей. Нередко в местах прикрепления связок и сухожилий, а также по краям суставных поверхностей формируются костные выросты – остеофиты. Прочность костей у пожилых людей значительно уменьшается и сравнительно небольшие травмы могут приводить к переломам.
Старение скелета характеризуется индивидуальной изменчивостью. У одних людей признаки старения появляются уже в 35–40-летнем возрасте, у других – только после 70 лет. В целом, признаки старения скелета у женщин выражены больше, чем у мужчин. Однако этот процесс существенно зависит от комплекса факторов: генетического, климатического, гормонального, алиментарного (фактор питания), функционального, экологического и т. д.
Как указывает В. С. Сперанский (1998), скелет человека – это совершенная динамическая конструкция, адаптированная к двигательной функции и образу жизни человека, чутко реагирующая на различные изменения, происходящие как в самом организме, так и в окружающей среде.
1.11. ВИДЫ ХРЯЩЕВОЙ ТКАНИ И ЕЕ СТРОЕНИЕ
Хрящевая ткань, как и костная, относится к скелетным тканям с опорно-механической функцией. По классификации выделяют три разновидности хрящевой ткани – гиалиновую, эластическую и волокнистую (рис. 1.6). Особенности строения различных видов хрящевой ткани зависят от места расположения ее в организме, механических условий, возраста индивидуума.
Наиболее широкое распространение у человека получила гиалиновая хрящевая ткань. Она входит в состав трахеи, некоторых хрящей гортани, крупных бронхов, темафизов костей, встречается в местах соединения ребер с грудиной и в некоторых других областях тела. Эластическая хрящевая ткань входит в состав ушной раковины, бронхов среднего калибра, некоторых хрящей гортани. Волокнистый хрящ обычно встречается в местах перехода сухожилий и связок в гиалиновый хрящ, например в составе межпозвоночных дисков.
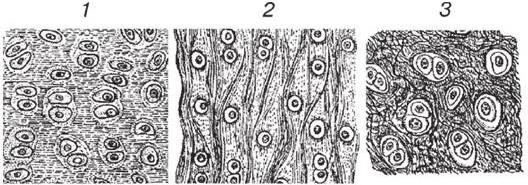
Рис. 1.6. Виды хрящевой ткани:
1 – гиалиновый хрящ; 2 – эластический хрящ; 3 – волокнистый хрящ
Строение всех видов хрящевой ткани в общих чертах сходно: они имеют в своем составе клетки и межклеточное вещество (матрикс). Одной из особенностей межклеточного вещества хрящевой ткани является его высокая обводненность: содержание воды в норме колеблется от 60 до 80 %. Площадь, занимаемая межклеточным веществом, значительно больше площади, занятой клетками. Межклеточное вещество хрящевой ткани вырабатывается клетками (хондробластами и молодыми хондроцитами) и имеет сложный химический состав. Оно подразделяется на основное аморфное вещество и фибриллярный компонент, который составляет примерно 40 % сухой массы межклеточного вещества и представлен в гиалиновой хрящевой ткани коллагеновыми фибриллами, образованными коллагеном II типа, идущими диффузно в различных направлениях. На гистологических препаратах фибриллы незаметны, так как имеют одинаковый с аморфным веществом показатель преломления. В эластической хрящевой ткани наряду с коллагеновыми фибриллами имеются многочисленные эластические волокна, состоящие из белка эластина, который тоже продуцируется хрящевыми клетками. Волокнистая хрящевая ткань содержит большое количество пучков коллагеновых волокон, состоящих из коллагенаIиIIтипа.
Ведущими химическими соединениями, образующими основное аморфное вещество хрящевых тканей (хондромукоид), являются сульфатированные гликозаминогликаны (кератосульфаты и хондроитинсульфаты А и С) и нейтральные мукополисахариды, большинство из которых представлено сложными надмолекулярными комплексами. В хрящах получили широкое распространение соединения молекул гиалуроновой кислоты с протеогликанами и со специфическими сульфатированными гликозаминогликанами. Этим обеспечиваются особые свойства хрящевых тканей – механическая прочность и в то же время проницаемость для органических соединений, воды и других веществ, необходимых для обеспечения жизнедеятельности клеточных элементов. Маркерными, наиболее специфичными для межклеточного вещества хряща соединениями являются кератосульфаты и определенные разновидности хондроитинсульфатов. Они составляют около 30 % сухой массы хряща.
Основные клетки хрящевой ткани – хондробласты и хондроциты. Хондробласты представляют собой молодые, малодифференцированные клетки. Они располагаются вблизи надхрящницы, лежат поодиночке и характеризуются округлой или овальной формой с неровными краями. Крупное ядро занимает значительную часть цитоплазмы. Среди клеточных органелл преобладают органеллы синтеза – рибосомы и полисомы, гранулярная эндоплазматическая сеть, комплекс Гольджи, митохондрии; характерны включения гликогена. При общегистологической окраске препаратов гематоксилином и эозином хондробласты слабобазофильны. Структура хондробластов указывает на то, что эти клетки обнаруживают высокую метаболическую активность, в частности, связанную с синтезом межклеточного вещества. Показано, что в хондробластах синтез коллагеновых и неколлагеновых белков пространственно разделен. Весь цикл синтеза и выведения высокомолекулярных компонентов межклеточного вещества в функционально активных хондробластах у человека занимает менее суток. Новообразованные белки, протеогликаны и гликозаминогликаны не располагаются непосредственно вблизи поверхности клетки, а распространяются диффузно на значительном расстоянии от клетки в образовавшемся ранее межклеточном веществе. Среди хондробластов встречаются и функционально неактивные клетки, строение которых характеризуется слабым развитием синтетического аппарата. Кроме того, часть хондробластов, находящаяся сразу под надхрящницей, не утратила способность к делению.
Хондроциты — зрелые клетки хрящевой ткани – занимают, главным образом, центральные участки хряща. Синтетические способности этих клеток значительно ниже, чем у хондробластов. Дифференцированные хондроциты чаще всего лежат в хрящевых тканях не поодиночке, а группами, включающими по 2, 4, 8 клеток. Это так называемые изогенные группы клеток, которые образовались в результате деления одной хрящевой клетки. Структура зрелых хондроцитов указывает на то, что они не способны к делению и заметному синтезу межклеточного вещества. Но некоторые исследователи считают, что при определенных условиях митотическая активность в этих клетках все же возможна. Функция хондроцитов заключается в поддержании на определенном уровне обменных метаболических процессов в хрящевых тканях.
Изогенные группы клеток находятся в хрящевых полостях, окруженных матриксом. Форма хрящевых клеток в изогенных группах может быть различной – округлой, овальной, веретеновидной, треугольной – в зависимости от положения на том или ином участке хряща. Хрящевые полости окружены узкой, более светлой, чем основное вещество, полоской, образующей как бы оболочку хрящевой полости. Эта оболочка, отличающаяся оксифильностью, называется клеточной территорией, или территориальным матриксом. Более удаленные участки межклеточного вещества называются интерстициальным матриксом. Территориальный и интерстициальный матриксы – участки межклеточного вещества с различными структурно-функциональными свойствами. В пределах территориального матрикса коллагеновые фибриллы ориентированы вокруг поверхности изогенных клеточных групп. Переплетения коллагеновых фибрилл образуют стенку лакун. Пространства между клетками внутри лакун заполнены протеогликанами. Интерстициальный матрикс характеризуется слабобазофильной или оксифильной окраской и соответствует наиболее старым участкам межклеточного вещества.
Таким образом, дефинитивная хрящевая ткань характеризуется строго поляризованным распределением клеток в зависимости от степени их дифференцировки. Вблизи надхрящницы находятся наименее дифференцированные клетки – хондробласты, имеющие вид вытянутых параллельно надхрящнице клеток. Они активно синтезируют межклеточное вещество и сохраняют митотическую способность. Чем ближе к центру хряща, тем клетки более дифференцированы, они располагаются изогенными группами и характеризуются резким снижением синтеза компонентов межклеточного вещества и отсутствием митотической активности.
В современной научной литературе описан еще один тип клеток хрящевой ткани — хондрокласты. Они встречаются только при разрушении хрящевой ткани, а в условиях ее нормальной жизнедеятельности не обнаруживаются. По своим размерам хондрокласты значительно крупнее, чем хондроциты и хондробласты, так как содержат в цитоплазме несколько ядер. Функция хондрокластов связана с активацией процессов дегенерации хряща и участием в фагоцитозе и лизисе фрагментов разрушенных хрящевых клеток и компонентов хрящевого матрикса. Иными словами, хондрокласты – это макрофаги хрящевой ткани, входящие в единую макрофагально-фагоцитарную систему организма.
1.12. ФИЗИОЛОГИЧЕСКАЯ И РЕПАРАТИВНАЯ РЕГЕНЕРАЦИЯ ХРЯЩЕВОЙ ТКАНИ
Ведущую роль в восстановительном процессе хрящевой ткани, как и любой другой ткани, играют закономерности ее эмбрионального развития (гистогенеза). Основные черты эмбрионального хондрогенеза обнаруживаются в ходе физиологической и репаративной регенерации. Следует особо подчеркнуть, что клеточные источники как физиологической, так и репаративной регенерации хрящевой ткани одни и те же.
В настоящее время не вызывает сомнения тот факт, что источником эмбрионального развития хрящевых тканей является мезенхима, в составе которой ведущую роль играют полипотентные мезенхимные стволовые клетки. Мезенхимным стволовым клеткам посвящен ряд исследований, из которых следует, что потомками этой клеточной популяции во взрослом организме человека являются стволовые стромальные клетки костного мозга. Они отличаются от стволовых кроветворных клеток, так как не экспрессируют антигены гемопоэтических клеток – CD34, CD14, CD45. Часть стволовых стромальных клеток при определенных условиях (соответствующее микроокружение, цитокиновое влияние, действие факторов роста и пр.) способна дифференцироваться по хондробластическому пути в хондрогенные клетки – прехондробласты, хондробласты, хондроциты. Предшественниками прехондробластов, по-видимому, являются малодифференцированные клетки, располагающиеся по ходу мелких кровеносных сосудов. Это так называемые периваскулярные клетки, которые иногда ошибочно называют перицитами. Термин «перицит» закреплен за зрелой, высокодифференцированной клеткой, лежащей в расщеплениях базальной мембраны кровеносных капилляров и осуществляющей связь между эндотелием и нервными элементами, вследствие чего может регулироваться просвет капилляра, вплоть до полного его закупоривания. Периваскулярные клетки часто еще именуют адвентициальными. В зависимости от условий они могут дифференцироваться в фибробласты, остеобласты, хондробласты и некоторые другие цитотипы. Важную роль в этом процессе играют факторы роста, представляющие собой малые полипептиды. Часть этих веществ является стимуляторами пролиферации прехондробластов и хондробластов. Например, AB- и BB-изоформы тромбоцитарного фактора роста, ТФР-â усиливают митотическую активность периваскулярных клеток и прехондробластов. Кислый и основной факторы роста фибробластов стимулируют пролиферацию, но подавляют дифференцировку хондробластов. Инсулиноподобные факторы роста1и2стимулируют и пролиферацию, и дифференцировку, причем последнюю – в гораздо большей степени. Действие факторов роста в различных сочетаниях является потенциальным механизмом для регуляции активности камбиальных клеток хрящевых тканей.
Если основным местом локализации стволовых клеток, необходимых для обеспечения физиологической и репаративной регенерации хрящевых тканей, является костный мозг, то главным источником камбиальных (активно делящихся) клеток служит соединительнотканная надхрящница. В ней (в отличие от хрящевой ткани) имеются кровеносные сосуды, и, следовательно, периваскулярные клетки. Последние, подвергаясь митотическому делению и дифференцировке, образуют популяцию прехондробластов, располагающихся во внутреннем слое надхрящницы и при благоприятных условиях микроокружения мигрирующих под надхрящницу и в самой хрящевой ткани превращающихся в хондробласты. Хондробласты активно пролиферируют и продуцируют межклеточное вещество. Через определенный промежуток времени эти клетки дифференцируются в хондроциты, формируя изогенные группы клеток.
Различают так называемый аппозиционный и интерстициальный рост хряща. Первый осуществляется за счет периваскулярных клеток и прехондробластов надхрящницы. Второй – за счет малодифференцированных хондробластов, лежащих в самой хрящевой ткани и находящихся непосредственно под надхрящницей.
Нужно иметь в виду, что полноценная репаративная регенерация хряща возможна лишь при небольших по площади повреждениях. При обширных повреждениях хрящевой ткани, сопровождающихся разрушением надхрящницы на большом протяжении, регенерацию хрящевой ткани опережает развитие грануляционной ткани на месте дефекта. С течением времени грануляционная ткань трансформируется в рубцовую соединительную ткань.
В последнее время активно развивается новое направление медицинской науки – регенеративная медицина, которая базируется на использовании новейших клеточных технологий, в частности тканевой инженерии. В передовых научных центрах разработаны методики выращивания в специальных условиях на особых питательных средах клеточных культур стволовых стромальных клеток и их производных, среди которых и предшественники хрящевых клеток. При необходимости эти клетки-предшественники могут быть пересажены (трансплантированы) в область дефекта хряща и служить источником его регенерации. К сожалению, пока работы в этом направлении носят преимущественно экспериментальный характер, но уже имеются сведения об использовании данной клеточной технологии в клинической практике.
1.13. РЕГУЛЯЦИЯ МЕТАБОЛИЧЕСКИХ ПРОЦЕССОВ В ХРЯЩЕВОЙ ТКАНИ
Метаболические процессы в хрящевой ткани регулируются многими механизмами. Один из ведущих регуляторных путей – эндокринный. Известно, что эстрогены и кортикостероидные гормоны тормозят или угнетают рост хрящей. Андрогены, гормоны щитовидной железы, соматотропный гормон (СТГ), наоборот, стимулируют рост и развитие хрящевой ткани, влияя на метаболизм и дифференцировку хрящевых клеток, способствуя оптимальному химическому составу межклеточного вещества.
Особая роль в регуляции гистофизиологии хрящевых тканей принадлежит цитокинам и факторам роста. Цитокины представляют собой полипептиды, которые продуцируются самыми различными клетками, в том числе и хрящевыми. Наибольший спектр биологически активных веществ, влияющих на метаболизм хрящевых тканей, выделяют макрофаги и тканевые базофилы (тучные клетки). Например, выделяемые ими фактор роста фибробластов, инсулиноподобный фактор роста, эпидермальный фактор роста оптимизируют соотношение процессов пролиферации и дифференцировки хрящевых клеток.
Активированные макрофаги выделяют ряд факторов, которые индуцируют процессы ангиогенеза при росте и регенерации хрящей. Различные монокины регулируют процессы пролиферации эндотелиальных клеток, их миграцию и новообразование капилляров надхрящницы независимо друг от друга. Тучные клетки выделяют также вазоактивные амины, способствующие повышению проницаемости сосудистой стенки, и фактор, стимулирующий рост капилляров. Выделяющиеся при дегрануляции тканевых базофилов вещества оказывают митогенное действие не только на эндотелиоциты, но и на другие клетки, в том числе находящиеся внутри хрящевой ткани.
Фактор роста эндотелия кровеносных сосудов, продуцируемый макрофагами и эндотелиоцитами, стимулирует развитие кровеносных сосудов микроциркуляторного русла надхрящницы, опосредованно оказывая благоприятное воздействие по трофику хряща.
Все факторы роста подразделяют на три группы. В первую входят стимуляторы пролиферативной активности клеток (митогены). Вторую группу составляют факторы, вызывающие пролиферацию и миграцию клеток хрящевой ткани. К третьей группе относятся модификаторы фенотипического состояния тканевых элементов.
Процесс старения сопровождается атрофией хрящевой ткани, изменением строения и количества клеток, изменением физико-химических свойств межклеточного вещества. Многие хрящевые клетки здесь гибнут, местами появляются довольно грубые волокнистые структуры, делающие основное вещество непрозрачным. В результате разжижения основного вещества появляются полости, в которые иногда проникают сосуды, и тут могут возникать островки окостенения. На участках стареющего гиалинового хряща могут отлагаться известковые соли, что приводит к обызвествлению хряща.
1.14. ВИДЫ СОЕДИНЕНИЙ КОСТЕЙ
Имеющиеся в теле человека многочисленные соединения костей целесообразно представить в виде классификации (табл. 1.1). В соответствии с данной классификацией существуют два основных вида соединений костей – непрерывное и прерывное, каждое из которых в свою очередь подразделяется на несколько групп (Гайворонский И. В., Ничипорук Г. И., 2005).
Следует отметить, что рельеф костей нередко отражает конкретный вид соединения. Для непрерывных соединений на костях характерны бугристости, гребни, линии, ямки и шероховатости, а для прерывных – гладкие суставные поверхности различной формы.
Непрерывные соединенияРазличают три группы непрерывных соединений костей – фиброзные, хрящевые и костные.
I. Фиброзные соединения, или соединения с помощью соединительной ткани, — синдесмозы. К ним относятся связки, мембраны, роднички, швы и вколачивания.
Связки – это соединения с помощью соединительной ткани, имеющие вид пучков коллагеновых и эластических волокон. По своему строению связки с преобладанием коллагеновых волокон называются фиброзными, а связки, содержащие преимущественно эластические волокна, – эластическими. В отличие от фиброзных, эластические связки способны укорачиваться и возвращаться к исходной форме после прекращения нагрузки.
Таблица 1.1
Виды соединений костей

По длине волокон связки могут быть длинными (задняя и передняя продольные связки позвоночного столба, надостистая связка), соединяющими несколько костей на большом протяжении, и короткими, соединяющими соседние кости (межостистые, межпоперечные связки и большинство связок костей конечностей).
По отношению к капсуле сустава различают внутрисуставные и внесуставные связки. Последние рассматривают как внекапсулярные и капсулярные. Связки как самостоятельный вид соединения костей могут выполнять различные функции:
– удерживающую или фиксирующую (крестцово-бугорная связка, крестцово-остистая, межостистые, межпоперечные связки и т. д.);
– роль мягкого скелета, так как являются местом начала и прикрепления мышц (большинство связок конечностей, связок позвоночного столба и т. д.);
– формообразующую, когда они вместе с костями формируют своды или отверстия для прохождения сосудов и нервов (верхняя поперечная связка лопатки, связки таза и т. д.).
Мембраны – это соединения с помощью соединительной ткани, имеющие вид межкостной перепонки, заполняющей в отличие от связок обширные промежутки между костями. Соединительнотканные волокна в составе мембран, преимущественно коллагеновые, располагаются в таком направлении, которое не препятствует движению. Роль их во многом сходна со связками. Они также удерживают кости относительно друг друга (межреберные мембраны, межкостные мембраны предплечья и голени), служат местом начала мышц (эти же мембраны) и формируют отверстия для прохождения сосудов и нервов (запирательная мембрана).
Роднички – это соединительнотканные образования с большим количеством промежуточного вещества и редко расположенными коллагеновыми волокнами. Роднички создают условия для смещения костей черепа в процессе родов и способствуют интенсивному росту костей после рождения. Наибольших размеров достигает передний родничок (30 × 25 мм). Он закрывается на втором году жизни. Задний родничок имеет размер 10 × 10 мм и полностью исчезает к концу второго месяца после рождения. Еще меньшие размеры имеют парные клиновидные и сосцевидные роднички. Они зарастают до рождения или в первые две недели после рождения. Роднички ликвидируются за счет разрастания костей черепа и формирования между ними шовной соединительной ткани.







