


Полная версия
Что ответить дарвинисту? Часть II
На подобную попытку сменить тему вестись не надо. Вы должны спокойно напомнить дарвинисту, что попросили его самого привести самые лучшие (на его взгляд) примеры наблюдаемой эволюции морфологических признаков вида под действием естественного отбора. А в ответ дарвинист привел Вам:
1) Примеры «эволюции» одного биологического вида… в этот же биологический вид. То есть, привел примеры колебаний внутривидовой изменчивости, уже известной для этого вида прежде.
2) Никакие строгие доказательства изменений именно в результате естественного отбора тоже не были представлены. Степень давления естественного отбора в конкретных природных популяциях не измерялась. Существенно ли хищническое давление крабов на популяции моллюсков? Или несущественно? Мы не знаем. А сами произошедшие изменения можно объяснить «вагоном и тележкой» других причин.
3) Дарвинист показал, что забрасывает оппонента ссылками, которые сам не читает.
4) Дарвинист показал, что забрасывает оппонента ссылками, не имеющими отношения к обсуждаемому вопросу.
5) Дарвинист показал, что склонен привирать (или ошибаться в свою пользу).
Вы должны пояснить дарвинисту, что последние три пункта вообще дискредитируют его, как оппонента. И на этом закончить.
Или, если есть еще желание, Вы можете пойти на поводу у дарвиниста еще один раз, и согласиться разобрать еще какое-нибудь второе (ну вот теперь уже точно самое убойное!) «доказательство эволюции», которое он горит желанием Вам изложить. Однако после этого второго захода – уже точно надо заканчивать.
2. Эволюционировавшие моллюски Андреевых
Раз уж мы так подробно разобрали «эволюцию американских улиток», то давайте тогда разберем еще и «эволюцию русских ракушек». Под «русскими ракушками» имеется в виду история с «эволюцией» двустворчатых моллюсков в Аральском море, которую описали в своей работе Андреевы (Андреева, Андреев, 2003). Этот пример «установленной эволюции» довольно популярен именно среди российских почитателей Дарвина. Большинству же зарубежных любителей эволюции этот пример, как правило, не знаком. Потому что все свои работы (посвященные разным аспектам биологии аральских моллюсков и других элементов экосистемы Аральского моря), Андреевы публиковали только в российских журналах. Соответственно, вычитали и разрекламировали этот пример «наблюдаемой эволюции» (вбросив его в широкие народные массы) тоже российские проповедники дарвинизма.
За многие годы исследований, Андреевы проделали очень большую работу, устанавливая те многочисленные изменения, которые происходили с экосистемами быстро высыхавшего Арала. По сути, это был протокол «умирания» целого моря.
К сожалению, благодаря стараниям верующих дарвинистов, широкую известность получила лишь самая слабая часть этих исследований – та глава, в которой авторы рассуждают именно об «эволюции» двустворчатых моллюсков Арала.[15]
Если пересказать эту главу совсем кратко, то авторы пишут, что в условиях быстро разрушающихся исконных экосистем Аральского моря, они обнаружили очень быструю эволюцию отдельных видов ракушек. Такую эволюцию продемонстрировали ракушки из родов Cerastoderma и Syndosmya. Причем продемонстрировали они её всего за 10–15 лет, что примерно равно такому же числу сменившихся поколений. То есть, и здесь тоже (снова) речь идет об «эволюционных» изменениях, происходивших с такой скоростью, которая кажется слишком быстрой для эволюции по дарвиновским механизмам.
Авторы работы описывают многочисленные изменения, происходившие в высыхающем море:
1) Постепенно возрастающую соленость.
2) Другие физико-химические изменения, вплоть до выпадения некоторых солей на дно в виде «корки».
3) Массовое вымирание прежде доминирующих видов.
4) Увеличение численности прежде редких видов, в том числе, завезенных в Арал из других водоемов.
5) Радикальную перестройку пищевых цепей Арала.
6) Массовую гибель макрофитов,[16] вплоть до того, что всё дно моря оказывалось устланным мертвыми остатками этих водных растений.
7) Расцветом фитопланктона.
Много внимания авторы уделяют постепенно возрастающей солености воды Аральского моря, что приводит к вымиранию сначала множества пресноводных видов, а затем и солоноватоводных, с заменой их чисто морскими и континентальными галофильными видами.
И вот в таких условиях конкретно для ракушки Cerastoderma (по утверждению авторов) наступаёт «звёздный час». Во-первых, эта ракушка демонстрирует взрыв внутривидовой изменчивости. То есть, начинает наблюдаться множество форм этого моллюска с изменениями по целому ряду морфологических признаков. Во-вторых, из моря исчезают основные «сдерживатели» популяций ракушки Cerastoderma – моллюски других родов, Dreissena и Hypanis. Hypanis являлся конкурентом Cerastoderma – так же как и Cerastoderma, он «сидел» практически целиком погруженным в грунт, и отфильтровывал взвешенные в воде питательные вещества. То есть, использовал ту же пищу, которой питалась и Cerastoderma. Таким образом, само существование Hypanis сдерживало численность Cerastoderma.
А ракушка Dreissena поступала с Cerastoderma еще жестче – стоило какой-нибудь Cerastoderma высунуть свою раковину из грунта, как на неё оседали личинки Dreissena и закреплялись на ней. Потому что дрейссенам для закрепления нужна какая-нибудь твердая основа. В ходе дальнейшего роста, дрейссены под тяжестью собственного веса постепенно «топили» Cerastoderma в грунте, попутно затрудняя им питание еще и за счет оплетания их своими прикрепляющими нитями (биссусом).[17]
Таким образом, в исконных экосистемах Арала распространение Cerastoderma сдерживалось доминированием дрейссен и Hypanis.
Однако в связи с повышением солености воды Аральского моря, дрейссены и Hypanis вымирают за относительно короткое время. А Cerastoderma не вымирают, потому что возросшая соленость воды пока еще их устраивает (находится в тех пределах, которые допустимы для этого вида). Вот тут-то Cerastoderma и показывает, на что она способна (в плане «эволюции»). Авторы работы отмечают, что в девяностых годах двадцатого века большинство особей Cerastoderma уже «вылезло» из грунта и находилось практически на его поверхности. Что было совсем не характерно для тех популяций Cerastoderma, которые существовали в Аральском море еще 20 лет назад. Авторы объясняют это тем, что в связи с вымиранием дрейссен, освобождается экологическая ниша, где можно питаться мелким планктоном, находясь прямо на поверхности грунта. При этом можно сколько угодно высовывать свою раковину над грунтом – никто уже не «садился» на эту раковину, и не оплетал её своими нитями.
Авторы работы приводят соответствующий рисунок, демонстрирующий это яркое изменение в пищевом поведении Cerastoderma (Рис. 8):
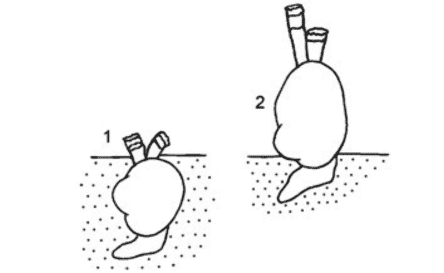
Рисунок 8. Схема положения Cerastoderma в грунте Аральского моря. 1 – типичное положение Cerastoderma isthmicum, 2 – положение современных Cerastoderma вне грунта. Из работы (Андреева, Андреев, 2003).
Помимо этого изменения в экологии, Cerastoderma 90-ых годов показывают еще и целый комплекс морфологических изменений, которые соответствуют их новому «имиджу» моллюсков, питающихся на поверхности грунта. А именно, у них смещается положение макушек раковины, меняется форма самой раковины (например, удлиняется её задний край), изменяется число и выраженность зубов и ребер раковины, изменяется строение замка (Андреева, Андреев, 2003). Авторы заключают, что большинство этих изменений биологически целесообразны. Они помогают моллюску удерживать устойчивое положение в новых для него условиях – на поверхности грунта. Кроме того, у большинства экземпляров наблюдается расширение мантийной линии в области сифонов.
Авторы приходят к выводу, что поскольку размах только что перечисленных морфологических изменений (т. е. установленных различий между Cerastoderma девяностых и семидесятых годов) выходит за пределы различий, характерных для разных видов двустворок (в других регионах), то здесь уже можно вести речь о факте видообразования. То есть, о появлении нового вида двустворчатого моллюска. Андреевы предполагают, что это видообразование произошло в результате стремительной эволюции под действием естественного отбора.
Такое же предположение авторы делают и в отношении установленных ими новых форм у других ракушек, из рода Syndosmya. Они заключают, что и эти морфологические варианты тоже возникли в результате именно эволюции моллюсков под соответствующим давлением естественного отбора (вследствие приспособления к грунтам разных типов).
Ну что же. Благодаря предыдущей главе, мы с Вами стали теперь уже опытными специалистами по «эволюции моллюсков», и можем понять, что в этом исследовании не так и почему говорить об «эволюции в ходе естественного отбора» здесь явно преждевременно. Давайте перечислим эти недостатки (в порядке убывающей важности):
1. Перед нами исследования, выполненные только одним коллективом авторов. Каких-либо независимых исследований (сделанных другими авторами), мы не имеем. По одной только этой причине говорить о строго установленном примере эволюции здесь уже вряд ли имеет смысл.
2. Самый главный недостаток конкретно этой работы – авторы не выяснили даже, являются ли обнаруженные ими изменения вообще наследственными? Или же установленные изменения ракушек имеют исключительно прижизненный характер? Может быть, все описанные изменения – это результат обыкновенной пластичности (модификационной изменчивости)? Действительно, весьма вероятно, что изменившееся поведение и морфология ракушек – простое следствие изменения условий среды. Может быть, конкретно этот вид ракушек всегда переходит к питанию на поверхности грунта, если в придонных слоях воды повышается количество пищи (планктона). Допустим, моллюск делает это, откликаясь именно на повышенное содержание планктона в соответствующих слоях воды. И при выходе на поверхность грунта у моллюска соответствующим образом изменяется еще и форма раковины. В этом случае мы будем наблюдать именно такую картину, которую наблюдали Андреевы – моллюски быстро и массово «вылезут» на поверхность грунта, демонстрируя при этом ряд соответствующих морфологических изменений… если в нужных слоях воды появится необходимое количество пищи. А оно там появилось, в связи с полным вымиранием других видов двустворок, которые раньше снижали содержание планктона в воде, а теперь снижать перестали (в связи с собственным вымиранием). Более того, авторы сами пишут об установленном ими повышении обилия планктона в воде Аральского моря.
То есть, возможно (и даже весьма вероятно), озвученные морфологические и поведенческие изменения, установленные авторами для этих видов ракушек, на самом деле, были, во-первых, характерны для этих видов. А во-вторых, являются не результатом эволюции, а просто результатом пластичности в ходе их индивидуального развития. В предыдущей главе мы видели, как улитки разных видов в ходе собственного роста образовывали более толстые раковины, если в воде присутствовал запах хищных крабов. Возможно, таким же образом и описанные ракушки Аральского моря способны выбираться из грунта, при этом соответствующим образом изменяя форму раковины, если в соответствующих слоях воды повышается обилие планктона? Или, допустим, если исчезают химические следы конкурента (Hypanis). Авторы работы не показали, что это не так.
Между тем, именно такое предположение наиболее вероятно. Чтобы его проверить, авторам необходимо было вырастить эти, «эволюционно измененные виды» ракушек в нормальных условиях (т. е. имитирующих условия Аральского моря в его исходном состоянии). И посмотреть, в кого конкретно превратятся изучаемые ими ракушки в таких условиях. Если эти ракушки в ходе роста всё равно продемонстрируют комплекс подмеченных авторами изменений, то значит, эти изменения, действительно, имеют наследственный характер. Если же в нормальных условиях «эволюционно измененные» ракушки тоже вырастут в совершенно нормальные формы (известные прежде), тогда это результат пластичности. И ни о какой «эволюции» в таком случае не может быть и речи.
Авторы такую проверку не сделали. Тем не менее, они почему-то пишут именно про «эволюцию моллюсков», игнорируя более простые варианты объяснений.
3. Авторы работы, скорее всего, справедливо связывают наблюдавшийся ими «взрыв» морфологической изменчивости моллюсков со стрессовыми условиями среды, в которые эти ракушки попали. Например, вот что они пишут по поводу развития личинок Syndosmya segmentum в воде Аральского моря:[18]
…Отставание в выклеве личинок в аральской воде по сравнению с контрольной азовской составляло 4 суток, а в оседании – 5 суток. Также было отмечено, что повышенное содержание иона Ca2+ приводит к появлению у личинок синдосмии уродливых недоразвитых раковинок (Карпевич, 1964).
Кроме того, авторы связывают взрыв изменчивости еще и с исчезновением стабилизирующего отбора в условиях вымирания основных конкурентов этих ракушек. Действительно, если раньше была серьезная борьба за выживание, навязанная конкурирующими видами, то выживать могли только такие ракушки, морфология которых была наиболее оптимальной для выживания. При исчезновении же основных конкурентов, выживать стало легче, и поэтому те морфологические варианты, которые раньше просто не доживали до того счастливого момента, когда их соберут в коллекцию биологи, теперь стали доживать, и удивлять биологов своим «взрывом разнообразия».
Однако непонятно, почему авторы работы пишут о повышении общего мутационного фона у этих ракушек:
Эволюционные изменения акклиматизированного в Аральском море двустворчатого моллюска синдосмия кратко можно представить следующим образом. Перемещение группы особей в водоем с водой, нарушающей ход эмбрионального развития, провоцирует взрыв мутагенеза.
Да, действительно, стрессовые условия вполне могут приводить к сбоям в индивидуальном развитии, приводящим к разнообразным (прижизненным) уродствам. Но почему авторы решили, что эти стрессовые условия (или вызванные ими уродства) каким-то образом должны повышать частоту мутаций у ракушек, и как следствие, приводить к повышенному разнообразию генотипов? Это предположение авторов не подтверждено ничем.
4. Наконец, вполне может быть и так, что установленные авторами различия между ракушками семидесятых и девяностых годов 20 века, действительно, являются наследственными. Но почему авторы решили, что за подобные изменения был ответственен именно естественный отбор? Авторы лишь предполагают, что какие-то из установленных ими форм ракушек – более приспособлены к одним условиям (например, к грунтам определенного типа), а какие-то формы более приспособлены к другим условиям. Однако авторы не попытались выяснить, а так ли это на самом деле. И даже если так, то на какую конкретно величину повышается приспособленность разных форм моллюсков в разных условиях? Ведь для того, чтобы всего за 10–15 поколений какая-то форма моллюсков (встречавшаяся раньше в единичных экземплярах) могла бы массово завоевать морское дно на соответствующих грунтах… для этого разница в общей выживаемости между разными формами ракушек должна составлять 200–300 %. Так ли это на самом деле?
Например, авторы пишут, что «вылезшие» на поверхность грунта ракушки Cerastoderma находились на этом грунте в довольно неустойчивом положении – когда подплывал аквалангист, ракушки схлопывали створки… и в результате сразу же падали на бок. Возникает вопрос, насколько безобидны такие «побочные эффекты» для моллюска, и перекрывают ли выгоды от питания именно на поверхности грунта столь неустойчивое положение раковины? А если перекрывают, то насколько конкретно? Особенно если учесть, что раньше эти же моллюски вполне успешно выживали, полностью погрузившись в грунт. И продолжают успешно выживать именно таким (погруженным) способом на участках, подверженных действию волн (Андреева, Андреев, 2003).
В общем, сила давления естественного отбора в данной работе не измерялась вообще никак. Просто было сделано предположение о том, что такая сила имелась, и что эта сила была достаточно мощной, чтобы отобрать соответствующие формы ракушек всего за 10–15 поколений. Понятно, что в качестве предположения, такие рассуждения, наверное, могут быть озвучены. Но вот о «строго установленном примере эволюции под действием естественного отбора» здесь говорить, конечно, не приходится.
5. Авторы работы пишут:
…Эта чрезвычайная скорость, с которой «притираются» возникшие фенотипы к новым условиям обитания, создает иллюзию направленного формирования фенотипов средой обитания. Однако, массовая ненаправленная изменчивость среди двустворок Арала (см. глава 6–7) в критические периоды свидетельствует о случайном характере процесса. И только массовое вовлечение в этот процесс двустворок, имеющих колоссальную энергию размножения, способствует успешной работе отбора. Именно массовость материала позволяет сохраняться и размножаться фенотипам (и генотипам), соответствующим новой среде обитания и постепенно (примерно за 10–15 лет), как в случае с эпифаунистическими Cerastoderma, выйти в новую адаптивную зону.
После прочтения этой цитаты напрашивается вопрос – почему авторы решили, что наблюдаемое ими «направленное формирование фенотипов средой обитания» – это именно «иллюзия», а не настоящая причина установленных изменений? То есть, если что-то «выглядит как утка, ходит как утка и крякает как утка»… то почему это всё-таки не утка, а «естественный отбор»?
Авторы объясняют (свой вывод о естественном отборе) наблюдаемой ими «массовой ненаправленной изменчивостью среди двустворок Арала в критические периоды». Но разве «массовая ненаправленная изменчивость» не могла быть просто массовыми сбоями индивидуального развития вследствие экологического стресса (т. е. просто уродствами), а вот наблюдаемое «выползание» ракушек на поверхность грунта (с соответствующими морфологическими изменениями раковин) – следствием именно «направленного формирования фенотипов средой обитания»? То есть, может быть, здесь нужно рассматривать мух отдельно, а котлеты – отдельно?
В свою очередь, «направленное формирование фенотипов средой обитания» – это, может быть, просто пример банальной экологической пластичности этих моллюсков?
Например, всем известно, что сосна в благоприятных условиях – это одно дерево, а вот на болоте – это уже совсем другое дерево. Ну а на обдуваемых ветром скалах – это уже третье дерево. Во всяком случае, морфологические различия между этими соснами будут никак не меньше, чем установленные различия между морфологией аральских ракушек. Так неужели из этого факта следует сделать вывод, что сосна таким образом… «эволюционирует под действием естественного отбора»? Нет, в данном случае сосна демонстрирует лишь тот размах экологической пластичности, к которой способен данный биологический вид. А эволюция здесь вообще ни при чем.
Далее авторы аргументируют свои рассуждения (о том, что это была именно эволюция естественным отбором) массовостью биологического материала, который предоставляют для эволюции обсуждаемые виды ракушек, благодаря их «колоссальной энергии размножения».
Но если так рассуждать, тогда куда более массовый «материал для эволюции» смогут легко предоставить, например, практически все виды планктонных водорослей. Или почти столь же колоссальные по численности и скорости сменяющихся поколений разнообразнейшие представители зоопланктона. Или даже многие виды рыб.[19] То есть, следуя подобной логике, специалисты, изучающие фитопланктон рек, озер и океанов, должны докладывать о новых фактах «свершившейся эволюции» чуть ли не каждый год – в каком-нибудь очередном полувысохшем озере, где плавают миллиарды особей каких-нибудь диатомовых водорослей, подвергаясь соответствующему экологическому стрессу. Или каких-нибудь коловраток. Или нематод. Или каких-нибудь кальмаров (в океане).
Но что-то мы не видим мощного потока научных публикаций, рапортующих нам о многочисленных фактах свершившейся (морфологической) эволюции. Наоборот, мы знаем, что даже самые массовые и быстро размножающиеся виды почему-то устойчиво сохраняют свой набор морфологических признаков именно в тех пределах, которые характерны для этих видов. Несмотря на многочисленные локальные изменения среды в самых разных регионах Земли. И вместо «мощного потока научных публикаций», нам приходится искать днем с огнём наши несчастные «двенадцать с половиной примеров эволюции» во всей живой природе. И один из таких примеров мы находим в книге Андреевых… в виде теоретических рассуждений о том, что обнаруженные ими изменения ракушек – это именно «эволюция под действием естественного отбора», потому что эти ракушки обладают «колоссальной энергией размножения». Что и говорить, просто «убойный» аргумент.
6. И наконец, как я уже говорил выше, установленные авторами изменения у аральских ракушек всё-таки могут быть наследственными. Такая возможность не исключена. И даже достаточно вероятна. Но эти изменения вполне могли быть обеспечены какими-нибудь другими механизмами, отнюдь не дарвиновскими. Например, под действием ныне модных эпигенетических механизмов.
Допустим, авторы этой работы просто натолкнулись на такие два вида ракушек, у которых имеется определенный набор генов, в обычных условиях «спящих». Но если эти гены проснутся, то они «переводят» данный вид ракушек в другой эколого-морфологический «режим» – из погруженного в грунт моллюска – в ракушку, питающуюся на поверхности грунта. Допустим, соответствующие гены «просыпаются» у моллюска под действием соответствующих химических веществ, растворенных в воде. Если эти вещества присутствуют в морской воде, то организм ракушки каким-то образом маркирует соответствующие гены (в половых клетках).
Какие-то гены включаются, какие-то, наоборот, выключаются.
Таким образом, программа индивидуального развития моллюска становится несколько другой, и в результате получается несколько другая ракушка, с несколько иным спектром морфологических и экологических признаков. И поскольку соответствующие гены уже «отмаркированы», то такие (отмаркированные) гены будут передаваться потомству.
То есть, соответствующие изменения морфологии и экологии станут наследственными. Таким образом, мы получим долговременную модификацию этого вида, которая будет устойчиво воспроизводиться в череде поколений, пока условия окружающей среды снова не вернутся к исходным.
Интересно, что характерными признаками подобного эпигенетического перехода должны являться, во-первых, быстрота этого перехода, а во-вторых, его массовость. То есть, сразу множество особей этих ракушек должны дружно продемонстрировать соответствующие изменения, причем это должно произойти быстро. Например, всего за 10–15 поколений. Потому что определенные химические вещества будут воздействовать на уже имеющиеся (соответствующие) гены сразу множества особей моллюсков. Таким образом, в случае подобного сценария генетических изменений, мы должны наблюдать именно такую картину «эволюции ракушек», которую и увидели реально авторы данной работы (массовые и быстрые изменения моллюсков).
В данном случае не представляется возможным как-то разделить, проверить и выбрать только один из двух возможных теоретических сценариев – «эволюцию вследствие естественного отбора» или же «эпигенетический переход между разными модификациями». Это можно сделать только в ходе дополнительных специальных исследований. А пока и «эволюция под действием естественного отбора», и «эпигенетический переход» являются вполне возможными сценариями. Действительно, эволюция за 10–15 поколений, хотя и с трудом, но всё же может быть обеспечена естественным отбором. Конечно, при условии, что соответствующее давление естественного отбора было очень сильным (если соответствующие признаки давали общее преимущество в выживании в пределах 200–300 %).