


Полная версия
Генетика человека с основами общей генетики. Учебное пособие
Столь гигантских полимеров, как ДНК, не выявлено больше ни в природе, ни среди искусственно синтезированных химических соединений. Длина молекулы ДНК первой хромосомы человека (самой крупной в наборе) достигает почти 8 см. Общая длина всех молекул ДНК клетки человека – около двух метров, а у саламандры почти в 30 раз больше.
Рибонуклеиновая кислота имеет множество разновидностей, но все ее молекулы построены по общим структурным принципам. Они состоят из одной полинуклеотидной цепочки, значительно более короткой, чем цепочка ДНК. В нуклеотидах РНК имеются 4 типа азотистых оснований: А, Г, Ц, У (урацил). РНК чаще, чем ДНК, содержит нетипичные нуклеотиды, которые обычно модифицируют ее функции. Углевод РНК – рибоза (С5Н10О5). Рассмотрим основные виды РНК в клетке.
Информационная (матричная) РНК – и-РНК (м-РНК). Содержит от нескольких сотен до десятков тысяч нуклеотидов. Молекула и-РНК представляет собой незамкнутую цепочку. Она переносит информацию о структуре белка с ДНК на рибосомы – место непосредственного синтеза полипептидной цепочки. У эукариот каждый белок клетки обычно кодируется отдельной молекулой и-РНК. У прокариот все гены одного оперона переписываются на одну общую молекулу и-РНК.
Рибосомальная РНК – р-РНК. Входит в состав рибосом. Помимо структурной функции, принимает непосредственное участие в синтезе полипептидной цепочки. Составляет 85 % всей РНК клетки. Прокариоты содержат 3 вида р-РНК, а эукариоты – 4 вида, весьма различных по размеру. Молекулы р-РНК и белков в субъединицах рибосом взаимодействуют упорядоченным образом.
Транспортная РНК – т-РНК. Переносит аминокислоты к месту синтеза белков на рибосомы. Каждая молекула т-РНК содержит немногим более 80 нуклеотидов. Специфичность т-РНК определяется структурой антикодона, т. е. участка соединения с определенным триплетом нуклеотидов и-РНК. Каждый антикодон определяет способность связываться с определенной аминокислотой на другом конце т-РНК. Эта способность зависит от активирующих ферментов, которые «узнают» соответствующие друг другу аминокислоты и т-РНК.
Гетерогенная ядерная РНК – гя-РНК. Является предшественником и-РНК у эукариот и превращается в и-РНК в результате сложных преобразований, которые будут рассмотрены в дальнейшем. Обычно гя-РНК значительно длиннее и-РНК.
Малая ядерная РНК – мя-РНК. Принимает участие в процессе преобразования гя-РНК.
РНК-праймер – крошечная РНК (обычно 10 нуклеотидов), участвующая в процессе репликации ДНК.
Для эволюционной биологии огромное значение имело выявление специфической каталитической активности некоторых РНК. Этот факт заставил многих ученых рассматривать РНК как «первомолекулу» в теориях происхождения жизни.
Нуклеиновые кислоты (ДНК и РНК) имеют характеристики первичной, вторичной и третичной структуры.
Первичная структура – последовательность нуклеотидов в полинуклеотидной цепочке.
Вторичная структура – порядок укладки полинуклеотидной нити.
Для ДНК вторичная структура – это двойная спираль нуклеотидных нитей. Существует несколько видов спиралей ДНК. Наиболее часто встречается правозакрученная спираль В-формы. Обнаружены участки ДНК, имеющие другую конфигурацию, как правозакрученную (А– и С-формы), так и левозакрученную (Z-форма).
РНК формирует вторичную конфигурацию за счет комплементарного соединения отдельных участков своей цепочки. Наиболее специфическую вторичную структуру имеет т-РНК (форма «клеверного листа»). Центральная петля молекулы т-РНК содержит антикодон. Очень сложную конфигурацию имеет вторичная структура р-РНК.
Третичная структура – различные виды компактизации молекулы нуклеиновой кислоты. В структуре ДНК это явление получило название суперспирализация. Третичная структура т-РНК похожа на букву «Г». Она меняется в зависимости от рН среды и других факторов. Особый случай представляет кольцевая ДНК (у бактерий, в митохондриях, в пластидах), образованная ковалентным соединением концов молекулы ДНК.
2.2. Репликация ДНК
Расшифровка структуры молекулы ДНК помогла объяснить принцип ее репликации. Репликацией называется процесс удвоения молекул ДНК. Этот процесс лежит в основе воспроизведения себе подобных живыми организмами, что является главным признаком жизни.
Особая роль ДНК в живом организме определяется такой ее фундаментальной особенностью, как способность к самоудвоению.
Гигантские молекулы ДНК эукариот имеют много участков репликации – репликонов, тогда как относительно небольшие кольцевые молекулы ДНК прокариот представляют каждая один репликон. Полирепликативный характер огромных молекул ДНК эукариот обеспечивает возможность ее репликации без одновременной деспирализации всей молекулы. Так, хромосомы клетки человека имеют более 50 000 репликонов, которые синтезируются как самостоятельные единицы. Если бы молекула ДНК эукариот удваивалась как один репликон, то этот процесс растянулся бы на несколько месяцев. Благодаря полирепликации он сокращается до 7–12 ч. В остальном в общих чертах процессы репликации прокариот и эукариот весьма похожи.
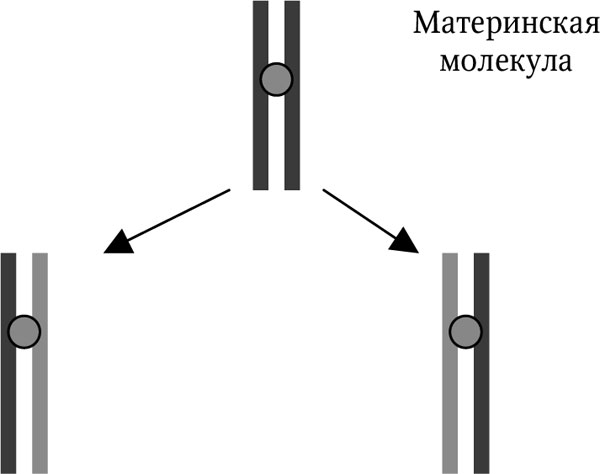
Рис. 2.2. Полуконсервативный принцип репликации ДНК
Процесс репликации ДНК в репликоне происходит в 3 этапа, в которых участвуют несколько разных ферментов.
Начинается репликация ДНК с локального участка, где двойная спираль ДНК (под действием ферментов ДНК-геликазы, ДНК-топоизомеразы и др.) раскручивается, водородные связи разрываются и цепи расходятся. В результате образуется структура, названная репликативной вилкой.
На втором этапе происходит типичный матричный синтез. К образовавшимся свободным связям присоединяются по принципу комплементарности (А-Т, Г-Ц) свободные нуклеотиды. Этот процесс идет вдоль всей молекулы ДНК. У каждой дочерней молекулы ДНК одна нить происходит от материнской молекулы, а другая является вновь синтезированной. Такая модель репликации получила название полуконсервативной (рис. 2.2). Этот этап осуществляет фермент ДНК-полимераза (известно несколько ее разновидностей).
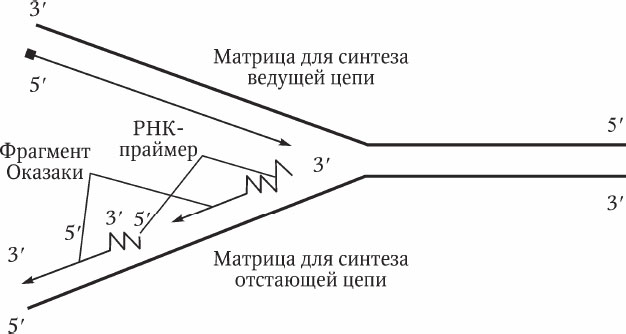
Рис. 2.3. Схема репликации ДНК
На двух материнских нитях синтез происходит неодинаково. Поскольку синтез возможен только в направлении 5' → 3', на одной нити идет быстрый синтез, а на другой – медленный, короткими фрагментами (1000–2000 нуклеотидов). В честь открывшего их биохимика Р. Оказаки они называются фрагментами Оказаки. Свободный 3'-конец, необходимый для начала синтеза фрагмента Оказаки, обеспечивает РНК-праймер, синтезируемая при помощи особой РНК-полимеразы – праймазы. После выполнения своей функции РНК-праймер удаляется, а ДНК-лигаза соединяет фрагменты Оказаки и восстанавливает первичную структуру ДНК (рис. 2.3).
На третьем этапе происходит закручивание спирали и восстановление вторичной структуры ДНК при помощи ДНК-гиразы.
Большинство ферментов, участвующих в репликации ДНК, работают в мультиэнзимном комплексе, связанном с ДНК. На основании этого американский биохимик Б. Альбертс выдвинул концепцию реплисомы, однако отдельные структуры, аналогичные рибосомам, пока не выявлены. Слаженная работа ферментов позволяет осуществлять репликацию с огромной скоростью: у прокариот – около 3000 п. н. (пар нуклеотидов) в секунду, у эукариот – 100–300 п. н. в секунду. Две новые молекулы ДНК представляют собой точные копии исходной молекулы.
Механизмы репликации весьма сложны, и многие детали этого процесса, особенно у высших животных, до настоящего времени неизвестны.
Глава 3. Цитогенетика
Наука не является и никогда не будет являться законченной книгой.
А. Эйнштейн (1879–1955), физик-теоретик, лауреат Нобелевской премии 1921 г.Цитогенетика – это раздел генетики, изучающий структурно-функциональную организацию генетического материала на уровне клетки, главным образом хромосом (Смирнов В. Н., 1990). Для всестороннего понимания организации генетического материала высших организмов (в том числе и человека) необходимы знания общих закономерностей упаковки ДНК во всех вариантах, предоставленных живой природой, – геномах вирусов, прокариот, протистов, клеточных органоидов.
3.1. Генетический материал вирусов и прокариот
Генетический материал вирусов представлен одной молекулой нуклеиновой кислоты (либо ДНК, либо РНК), окруженной защитной белковой оболочкой – капсидом. Функционирование вирусов происходит по-разному, в зависимости от их свойств и структуры, но всегда с помощью ферментативной системы клетки-хозяина. Вирусы могут существовать только как внутриклеточные паразиты. До сих пор не закончен давний научный спор, можно ли считать вирус живым: «существо или вещество».
Существуют вирусы, имеющие одно– и двухцепочечные РНК, и вирусы, имеющие одно– и двухцепочечные ДНК, причем обе группы ДНК-содержащих вирусов имеют представителей с линейными и кольцевыми формами. У аденовирусов двухцепочечная ДНК связана с терминальным белком, а у вируса оспы ДНК замкнута на концах ковалентной связью (Льюин Б., 1987).
РНК-содержащие вирусы более разнообразны. Так, выделяют вирусы с «плюс-цепью», которые сразу могут функционировать, и вирусыс «минус-цепью», которые вначале должны построить «плюс-цепь» с помощью РНК-полимеразы клетки-хозяина. Двухцепочечные вирусы представляют собой варианты соединенных цепей без расхождения после синтеза второй цепи. Особую группу РНК-содержащих вирусов составляют ретровирусы, которые будут рассмотрены ниже. Размеры РНК-содержащих вирусов обычно варьируют в пределах 3000–7000 нуклеотидов, а самый маленький из них имеет всего 1200 рибонуклеотидов и 1 структурный ген, кодирующий белок оболочки капсида.
ДНК-содержащие вирусы, особенно фаги (вирусы бактерий), обычно значительно крупнее РНК-содержащих. Так ДНК фага Т4 содержит 180 000 п. н. и кодирует множество белков. Крупные молекулы ДНК вирусов компактно упакованы внутри капсида благодаря суперспирализации.
Возможны два варианта развития вируса в клетке: либо интеграция с геномом хозяина – лизогения, либо синтез вирусных частиц на основе генетической программы вируса, но с помощью метаболической системы хозяина – лизис. Второй вариант обычно приводит к разрушению клетки-хозяина. Факт регуляции генной активности вируса, его способности существовать в интегрированной форме, был доказан в работах нобелевского лауреата 1965 г., французского микробиолога А. Львова (1902–1994). Интегрированная форма вируса получила название профаг. Под действием внешних факторов (например, УФ-облучение) возможна активация профага и вновь превращение его в фаг.
Вирусы обычно обладают специфичностью в отношении клеток организма хозяина.
Геном прокариот представлен одной кольцевой молекулой ДНК, формирующей компактную структуру нуклеоида посредством суперспирализации. Весьма хорошо изучен геном кишечной палочки (Escherichia coli) – классического генетического объекта, у которой идентифицировано более 4200 генов. ДНК E. coli содержит 4,6 млн п. н. Наименьший размер генетического материала у живых организмов (не будем относить к ним вирусы) отмечен у микоплазмы: 600 000 п. н. и около 500 генов. Эти данные и послужили основой для теоретических расчетов, которые показали, что элементарная «машина жизни» может работать при наличии всего 350 генов.
Главная особенность организации генома прокариот – это их объединение в группы, или кластеры, с общей регуляцией. Группа структурных генов прокариот, находящихся под контролем одного регуляторного участка, называется опероном (Miller J., Reznikoff W., 1978). Организация генетического материала по типу оперона позволяет бактериям быстро переключать метаболизм с одного субстрата на другой. Бактерии не синтезируют ферменты определенного метаболического пути в отсутствие необходимого субстрата, но способны в любой момент начать их синтез при появлении этого субстрата. Структура и функционирование оперона были показаны в работах знаменитых французских биохимиков Ж. Моно (1910–1976) и Ф. Жакоба, разделивших с А. Львовым Нобелевскую премию 1965 г. Регуляцию по типу оперона мы рассмотрим ниже.
Особый интерес представляют плазмиды – небольшие кольцевые молекулы ДНК внутри бактериальной клетки. Подобно вирусам, плазмиды способны либо интегрироваться с бактериальной ДНК, либо существовать обособленно от нее. Крупные плазмиды присутствуют в клетке в количестве 1–3 копий, мелкие могут быть представлены десятками копий. Хорошо изучена самая первая из обнаруженных плазмид, крупная плазмида F бактерии E. coli. Она представляет собой кольцевую молекулу ДНК величиной в 100 тыс. п. н. и содержит более 60 генов. Плазмида F обеспечивает содержащим ее бактериальным клеткам возможность взаимодействовать с бесплазмидными бактериями и передавать им свою генетическую информацию.
Многие авторы считают, что плазмиды являются одной из разновидностей вирусов и между ними нет принципиальных различий (Жданов В. М., 1988; Кусакин О. Г., Дроздов А. Л., 1994).
3.2. Генетический материал эукариот
Генетический материал эукариот сконцентрирован в ядре и представлен хромосомами, в которых молекула ДНК образует сложный комплекс с различными белками.
Каждая клетка любого организма содержит определенный набор хромосом. Совокупность хромосом клетки называется кариотипом (рис. 3.1). Количество хромосом в клетке не зависит от уровня организации живых организмов – некоторые протисты имеют их более тысячи. У человека в кариотипе 46 хромосом, у шимпанзе – 48, у крысы – 42, у собаки – 78, у коровы – 60, у дрозофилы – 8, у тутового шелкопряда – 56, у картофеля – 48, у рака-отшельника – 254 и т. д.
В кариотипе соматических клеток выделяются пары одинаковых (по форме и генному составу) хромосом – так называемые гомологичные хромосомы (1-я – материнская, 2-я – отцовская). Набор хромосом, содержащий пары гомологов, называется диплоидным (обозначается 2n). Половые клетки – гаметы, содержат половину диплоидного набора, по одной хромосоме из каждой пары гомологов. Такой набор называется гаплоидным (обозначается n).

Рис. 3.1. Кариотип человека
Исследуется кариотип обычно на стадии метафазы митоза, когда каждая хромосома состоит из двух идентичных хроматид и максимально спирализована. Соединяются хроматиды в области центромеры (первичной перетяжки). В этой области при делении клетки на каждой сестринской хроматиде образуется фибриллярное тельце – кинетохор, к которому присоединяются нити веретена деления.
Концевые участки хромосом получили название теломеры. Они препятствуют слипанию хромосом, т. е. ответственны за их «индивидуальность». Теломеры имеют специфический состав ДНК, связанной со специфическим комплексом белков. Состав теломерной ДНК весьма «консервативен» у разных видов. В последние годы теломеры привлекают к себе внимание в связи с проблемой старения клеток и долголетия. Дело в том, что у взрослого организма с каждым новым делением клетки теряется участок теломеры. Потеря всей теломеры приводит к смерти клетки. Понимание генетического контроля этого явления поможет решить многие проблемы медицины.
Участок хроматиды между центромерой и теломерой называется плечом. Плечи имеют свои обозначения: короткое – р и длинное – q. В зависимости от расположения центромеры различают следующие морфологические типы хромосом:
– метацентрические (p = q);
– субметацентрические (q > p);
– акроцентрические (одноплечие – q).
Такое морфологическое разнообразие характерно для большинства организмов. К нему добавляется разнообразие хромосом по размерам. Не совсем понятен биологический смысл этого явления. Известно, что хромосомы – это не просто «кладовые» генетической информации, а активно функционирующие структуры. Их основная биологическая роль заключается в обеспечении равномерности распределения генетического материала при делении клетки и рекомбинации при мейозе. Возможно, морфологическое разнообразие способствует более успешному выполнению этой роли (Гринев В. В., 2006). Хотя можно отметить, что у одних животных хромосомы морфологически удивительно однообразны, хотя и различаются по размерам (лошадь, корова), у других – разнообразны(человек).
Конец ознакомительного фрагмента.
Текст предоставлен ООО «ЛитРес».
Прочитайте эту книгу целиком, купив полную легальную версию на ЛитРес.
Безопасно оплатить книгу можно банковской картой Visa, MasterCard, Maestro, со счета мобильного телефона, с платежного терминала, в салоне МТС или Связной, через PayPal, WebMoney, Яндекс.Деньги, QIWI Кошелек, бонусными картами или другим удобным Вам способом.


