


Полная версия
Медицинская микробиология, иммунология и вирусология
Таким образом, в классификации Берги-2001 (George M. Garrity, Julia A. Bell, Timothy G. Lilburn. Taxonomic Outline of the Prokaryotes. Bergey’s Manual of Systematic Bacteriology, Second Edition. Release 5.0, May 2004) выделены и охарактеризованы 2 домена, 26 типов, 42 класса и большое количество семейств и родов бактерий.
В определителе Берги дано следующее определение бактерий (прокариот): «единичные клетки или простые скопления сходных клеток размером 0,2 × × 10,0 мкм, которые образуют своеобразные групповые структуры. Ядерный аппарат (нуклеоид, или генофор) никогда не отделен от цитоплазмы системой унитарных (элементарных) мембран. Клеточное деление не связано с циклическими изменениями строения клетки или изменением окрашиваемости ядерного аппарата или цитоплазмы; система микротрубочек (нитей веретена) не образуется. Цитоплазматическая мембрана обычно представляет собой топологический комплекс (элементарная мембрана) и образует ячеистые, ламеллярные (пластинчатые) или трубчатые впячивания в цитоплазму; вакуоли и репликативные цитоплазматические органеллы не связаны с системой плазматической мембраны, встречаются относительно редко (газовые вакуоли; хлоробиум-везикулы, т. е. пузырьки, окруженные однослойной мембраной и содержащие аппарат фотосинтеза у некоторых фотобактерий) и окружены неунитарными мембранами. Дыхательные и фотосинтетические функции связаны с системой плазматической мембраны у тех организмов, которые обладают этими физиологическими функциями, хотя у цианобактерий они могут быть и не связаны с плазматической и тилакоидными (тилакоиды – двойные ламеллы) мембранами. Рибосомы типа 70S, кроме одной группы бактерий – архебактерий, у которых они имеют более высокое значение S и распределены по цитоплазме; эндоплазматического ретикулума с прикрепленными рибосомами нет. Цитоплазма неподвижна, ее переливания, образования псевдоподий, эндо- и экзоцитоза не происходит. Питательные вещества потребляются в молекулярной форме. Клетки окружены ригидной стенкой, хотя она имеется не у всех бактерий (ее нет у микоплазм и некоторых архебактерий). Клетки могут быть неподвижными или могут обладать плавательной подвижностью, обеспечиваемой жгутиками бактериального типа, или скользящей подвижностью на плотной поверхности. Прокариоты – преимущественно одноклеточные организмы, однако образование нитевидных мицелиальных и колониальных форм также происходит. Они обладают механизмами переноса генов и рекомбинации, но эти процессы происходят без образования гамет (половых клеток) и зигот».
Основные признаки, по которым дифференцируют прокариот и эукариот, приведены в табл. 1.
Для обозначения видов бактерий используют бинарную номенклатуру, состоящую из названия рода (пишется с заглавной буквы) и вида (пишется всегда со строчной буквы и состоит из одного слова), например, Shigella flexneri (возбудитель дизентерии – род Shigella, вид flexneri). Когда название вида неоднократно повторяется, то первый раз название рода пишется полностью, а затем пишется только начальная буква его. Например, Shigella flexneri – S. flexneri. В связи со сложностью классификации бактерий названия внутривидовых форм (подвидов, серотипов и подсеротипов) часто используются как видовые, т. е. в качестве второго слова биномена – Salmonella enteritica (видовое), S. indica (подвидовое), S. typhimurium (серотип).
Таблица 1
Некоторые дифференциальные признаки прокариот и эукариот1 (Bergey’s Manual of Determinative Bacteriology–9)
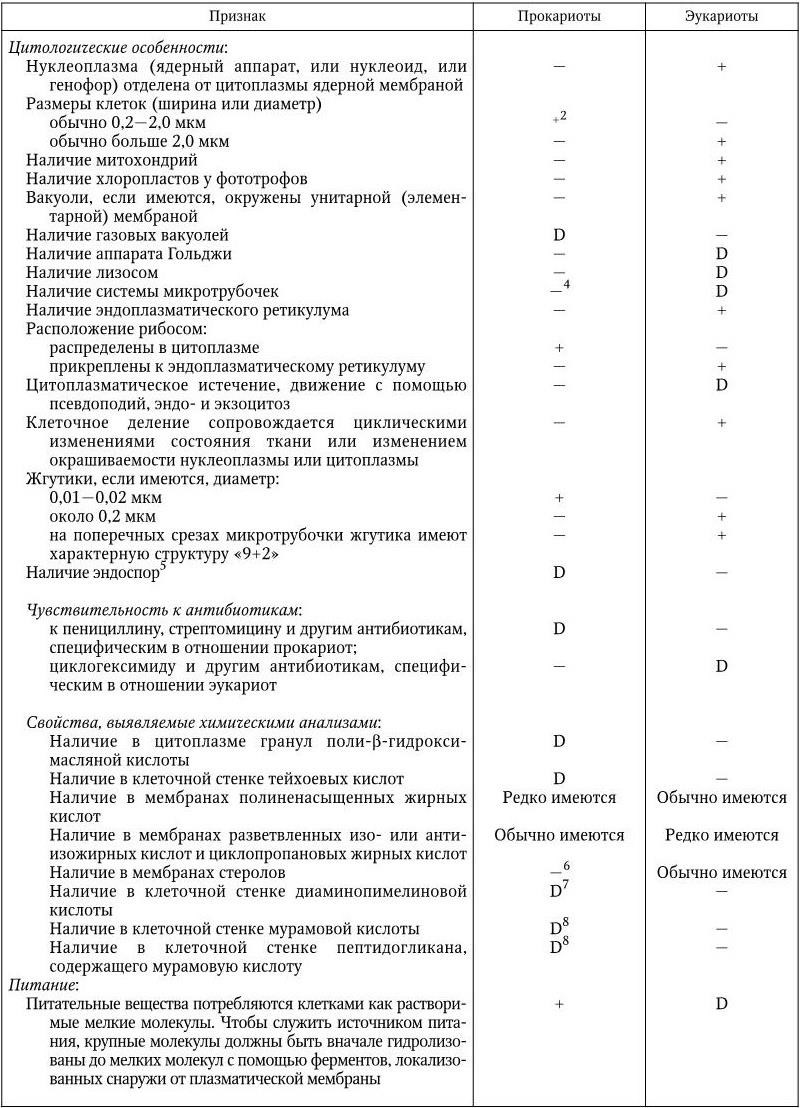
2 Некоторые бактерии (например, определенные трепонемы, микоплазмы, Haemobartonella) могут иметь ширину меньше 0,1 мкм, а другие бактерии (например, Achromatium, Macromonas) – больше 10 мкм.
3 Газовые вакуоли не ограничиваются элементарной мембраной. Везикулы, составляющие вакуоль, могут подвергаться коллапсу при внезапном гидростатическом воздействии – свойство, существенное для их идентификации.
4 Некоторые внутриклеточные фибриллы, которые могут быть микротрубочками, были описаны у Spiroplasma, цианобактерий Anabaena, некоторых спирохет и у L-форм бактерий.
5 Эндоспоры бактерий обычно устойчивы при температуре 80 °C или выше в течение 10 мин. Однако некоторые эндоспоры могут погибать при такой температурной обработке и должны быть испытаны при более низкой температуре.
6 Кроме мембран большинства микоплазм.
7 Имеются у всех грамотрицательных эубактерий и у многих грамположительных.
8 Имеются в клеточной стенке всех эубактерий, кроме хламидий; отсутствуют у архебактерий.
9 У цианобактерий они не связаны с цитоплазматической и тилакоидными мембранами.
10 За редкими исключениями, например у некоторых фотобактерий.
11 Кроме митохондрий, в которых встречается тип Mn.
12 Кроме митохондрий и хлоропластов, которые имеют 70S рибосомы.
Вопрос о самозарождении и развитии жизни на Земле
Вопрос о самозарождении и развитии жизни на Земле был и остается одним из самых главных и самых трудных вопросов науки. Теперь уже ни у кого нет сомнения в том, что самозарождение жизни могло происходить лишь после того, как возникнут чисто химическим путем важнейшие органические соединения, необходимые для того, чтобы произошел синтез прежде всего первородных генов, т. е. генов, образующихся без участия белков, до их возникновения; первородных белков, т. е. белков, которые образуются без участия генов, и генетического кода, так как без него ген не может реализовать свою задачу. В самом деле, синтез генов у всех живых организмов происходит только при участии сложной системы биосинтеза ДНК, а синтез белков происходит только по программе, заключенной в структуре гена: порядок расположения кодонов в гене определяет порядок расположения аминокислот в белке. Вот почему и возник вопрос: что возникло раньше – ген или белок? Образно говоря, что возникло раньше – курица или яйцо?
Выдающийся русский ученый А. И. Опарин, который внес большой вклад в развитие так называемой коацерватной теории происхождения жизни, получившей в XX в. общее признание, назвал этот вопрос чисто схоластическим. Однако он ошибся. Изучение структуры гена и генетического кода не оставляет никаких сомнений в том, что генетической системе принадлежит важнейшая роль в самозарождении и эволюции жизни на Земле. Нет более никакого сомнения в том, что именно ген служит основным носителем и хранителем жизни на Земле, а белок – ее творцом, поэтому вопрос о том, как возникли первородные гены, первородные белки и генетический код, приобрел основное значение для выяснения механизма зарождения жизни. Следует при этом иметь в виду, что структуры, состоящие только из первородных генов и первородных белков, сами по себе еще не способны к самостоятельному размножению, как это хорошо демонстрируют простейшие живые организмы – плазмиды и вирусы. Для того чтобы процесс самозарождения жизни состоялся, необходимо было возникновение специализированных систем жизнеобеспечения. К ним относятся следующие системы:
1. Система биологического самовоспроизводства генов, т. е. система биосинтеза ДНК.
2. Сложная биологическая система синтеза белков, которая включает в себя целый комплекс различных компонентов (мРНК, тРНК, рибосомы и комплекс особых рабочих белков).
3. Система мобилизации энергии, необходимой для синтеза всех компонентов формирующейся первородной клетки.
4. Система мембран, с помощью которых формирующаяся клетка отграничивается от внешней среды, сохраняя способность осуществлять активную и пассивную связь с ней.
5. Система, обеспечивающая саморегуляцию выражения генетической информации.
6. Система саморегуляции клеточного деления, т. е. размножения клетки.
Только после формирования всех этих систем жизнеобеспечения и возникновения уникальной структурной единицы живой материи – клетки – завершается этот первый и важнейший этап самозарождения и самоутверждения жизни на Земле. Эти вопросы более подробно рассматриваются в главе 74. Последующие этапы эволюции включали в себя появление многоклеточных организмов и их дальнейшую эволюцию в направлении растительного и животного царств. Изучением генетических механизмов эволюции занимается специальная наука – эволюционная генетика, или геномика. Однако нельзя не обратить особое внимание на предлагаемую представителями геномики гипотезу, получившую название пульсации генома. Суть ее состоит в том, что изменение генома может идти не только в сторону нарастания количества генов, но и в сторону его уменьшения. Предполагается, что это определяется не чем иным, как полинуклеотидным выбором (Пн-выбором) ДНК-реципиента. Из этого следует, что предметом естественного отбора служит не фенотипический признак, кодируемый донорной ДНК, а новые последовательности ДНК, независимо от того, какие признаки они кодируют. С этих позиций геномики естественный отбор складывается из двух этапов: Пн-выбора и фенотипического дарвиновского отбора. Такой вывод полностью совпадает с утверждением о том, что ген служит главным носителем и хранителем жизни, ее главным архитектором, т. е. именно ген играет важнейшую роль в эволюции самой живой материи.
Глава 4
Морфология бактерий
Формы бактерий
Всем бактериям присущи определенная форма и размеры, которые выражаются в микрометрах (мкм). Они варьируют в широких пределах – от 0,1 – 0,15 (Mycoplasma) до 10 – 15 мкм (Clostridium) в длине и от 0,1 мкм до 1,5 – 2,5 мкм в диаметре. Бо́льшая часть бактерий имеет размеры 0,5 – 0,8 мкм × 2 – 3 мкм.
Различают следующие основные формы бактерий: шаровидные (сферические), или кокковидные (греч. kokkos – зерно); палочковидные (цилиндрические); извитые (спиралевидные); нитевидные. Кроме того, существуют бактерии, имеющие треугольную, звездообразную, тарелкообразную форму (см. цв. вкл., рис. 1.1 – 1.8). Обнаружены так называемые квадратные бактерии, которые образуют скопления из 8 или 16 клеток в виде пласта (рис. 1.7).
Кокковидные патогенные бактерии обычно имеют форму правильного шара диаметром 1,0 – 1,5 мкм; некоторые – бобовидную, ланцетовидную, эллипсоидную форму. По характеру взаиморасположения образующихся после деления клеток кокки подразделяют на следующие группы:
1. Микрококки (греч. mikros – малый). Делятся в одной плоскости, располагаются одиночно и беспорядочно; сапрофиты; патогенных для человека нет (рис. 1.1).
2. Диплококки (греч. diplos – двойной). Деление происходит в одной плоскости с образованием пар клеток, имеющих либо бобовидную (Neisseria gonorrhoeae), либо ланцетовидную (Streptococcus pneumoniae) форму (рис. 1.2).
3. Стрептококки (греч. streptos – цепочка). Деление клеток происходит в одной плоскости, но размножающиеся клетки сохраняют между собой связь и образуют различной длины цепочки, напоминающие нити бус. Многие стрептококки являются патогенными для человека и вызывают различные заболевания: скарлатину, ангину, гнойные воспаления и др. (рис. 1.3).
4. Стафилококки (греч. staphyle – гроздь винограда). Деление происходит в нескольких плоскостях, а образующиеся клетки располагаются скоплениями, напоминающими гроздья винограда. Стафилококки вызывают более 100 различных заболеваний человека. Они наиболее частые возбудители гнойных воспалений (рис. 1.4).
5. Тетракокки (греч. tetra – четыре). Деление клеток происходит в двух взаимно перпендикулярных плоскостях с образованием тетрад. Патогенные для человека виды встречаются очень редко (рис. 1.5).
6. Сарцины (лат. sarcina – связка, тюк). Деление клеток происходит в трех взаимно перпендикулярных плоскостях с образованием пакетов (тюков) из 8, 16, 32 и большего числа особей. Особенно часто встречаются в воздухе. Имеются условнопатогенные представители (рис. 1.6).
Палочковидные (цилиндрические) формы бактерий. Термин «бактерия» (греч. bakterion – палочка) применяется как для названия всего царства прокариот (Eubacteria, Archebacteria), так и для названия палочек, не образующих спор. Палочки, образующие споры, подразделяют на бациллы (лат. bacillus – палочка) – аэробные спорообразующие бактерии, например Bacillus anthracis – возбудитель сибирской язвы, и клостридии (лат. clostridium – веретенообразный) – анаэробные спорообразующие бактерии, например Clostridium tetani – возбудитель столбняка. Палочки бывают длинными – более 3 мкм (Clostridium novyi – возбудитель газовой гангрены), короткими – 1,5 – 3,0 мкм (Escherichia coli и большинство возбудителей кишечных инфекций) и очень короткими – менее 1,0 мкм – в виде коккобактерий (Francisella tularensis – возбудитель туляремии, Brucella melitensis – бруцеллеза). Концы палочек могут быть закругленными (Escherichia coli и др.), заостренными (Fusobacterium), утолщенными (Corynebacterium), обрезанными (Bacillus anthracis);
палочка может иметь овоидную (яйцевидную) форму (Yersinia pestis – возбудитель чумы). По диаметру их делят на тонкие (Mycobacterium tuberculosis – возбудитель туберкулеза) и толстые (Clostridium perfringens – возбудитель газовой гангрены). По взаиморасположению бактерий их подразделяют на три группы (см. цв. вкл., рис. 2.1 – 2.6): 1) монобактерии – палочки располагаются одиночно и беспорядочно, сюда относится большинство палочковидных форм (рис. 2.1); 2) диплобактерии, располагающиеся попарно (Pseudomonas) (рис. 2.2); 3) стрептобактерии (Haemophilus ducreyi – возбудитель мягкого шанкра) или стрептобациллы (Bacillus anthracis) – бактерии, располагающиеся цепочкой (рис. 2.3 и 2.4).
Извитые (спиралевидные) бактерии по количеству и характеру завитков, а также по диаметру клеток подразделяют на две группы: 1) вибрионы (греч. vibrio – извиваюсь, изгибаюсь) имеют один изгиб, не превышающий четверти оборота спирали, однако могут иметь и форму прямой палочки, без изгиба (Vibrio cholerae – возбудитель холеры) (рис. 2.5); 2) спириллы (греч. speira – спираль) – клетки, имеющие большой диаметр и малое (2 – 3) число завитков (Spirillum minor – возбудитель содоку) (рис. 2.6). Особую группу спиралевидных бактерий представляют спирохеты, выделенные в порядок Spirochaetales. Их морфология подробно описана в гл. 69.
Нитевидные формы бактерий. Различают два типа нитевидных бактерий: образующие временные нити и постоянные.
Временные нити, иногда с ветвлениями, образуют палочковидные бактерии при нарушении условий их роста или регуляции клеточного деления (микобактерии, коринебактерии, а также риккетсии, микоплазмы, многие грамотрицательные и грамположительные бактерии). При восстановлении механизма регуляции деления и нормальных условий роста эти бактерии восстанавливают обычные для них размеры.
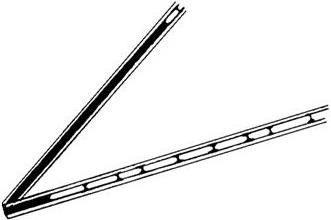
Рис. 2.7. Нитевидные бактерии.
Sphaerotilus natans, часть влагалища пустая
Постоянные нитевидные формы образуются из палочковидных клеток, соединяющихся в длинные цепочки либо с помощью слизи, либо чехлами (влагалищами, рис. 2.7), либо мостиками. Влагалищами, или футлярами, называют трубковидные чехлы гетерополисахаридной природы. Слизь может связывать отдельные клетки в длинные нити (Zoogloea) или пленки (Bacteriogloea). Нитевидные формы образуют серобактерии и железобактерии.
Следует особо отметить, что бактерии отличаются высоким полиморфизмом (индивидуальной изменчивостью формы, не передающейся по наследству), особенно при культивировании на искусственных питательных средах. Под действием различных факторов (антибиотиков, химических веществ) могут возникать необычные по форме и величине клетки, которые, однако, способны ревертировать в исходное состояние при снятии действия этих факторов.
Строение бактериальной клетки
Клетка – универсальная структурная единица живой материи. Подтверждением этому является сходство в химическом составе бактерии и клетки млекопитающего, которая в 2000 раз больше первой (табл. 2).
Таблица 2
Примерный химический состав типичной бактерии и типичной клетки
Организация бактериальной клетки позволяет ей координировать все процессы жизнедеятельности, за определенный срок удваивать свою биомассу и размножаться путем бинарного деления. В составе бактериальной клетки можно выделить различные структуры (рис. 3):
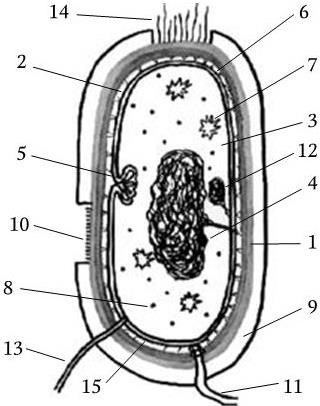
Рис. 3. Схема строения бактериальной клетки:
1 – клеточная стенка; 2 – цитоплазматическая мембрана; 3 – цитоплазма; 4 – нуклеоид; 5 – мезосома; 6 – периплазматическое пространство; 7 – включения; 8 – рибосома; 9 – капсула; 10 – микрокапсула; 11 – жгутик; 12 – плазмида; 13 – донорная ворсинка; 14 – фимбрии (реснички); 15 – перемычки в периплазматическом пространстве
Клеточная стенка
Клеточная стенка – структурный компонент, присущий только бактериям (кроме микоплазм). Клеточная стенка выполняет следующие функции:
1. Определяет и сохраняет постоянную форму клетки.
2. Защищает внутреннюю часть клетки от действия механических и осмотических сил внешней среды.
3. Участвует в регуляции роста и деления клеток.
4. Обеспечивает коммуникацию с внешней средой через каналы и поры.
5. Несет на себе специфические рецепторы для бактериофагов.
6. Определяет во многом антигенную характеристику бактерий (природу и специфичность О- и К-антигенов).
7. Содержащийся в ее составе пептидогликан наделяет клетку важными иммунобиологическими свойствами (см. ниже).
8. Нарушение синтеза клеточной стенки бактерий является главной причиной их L-трансформации.
Строение клеточной стенки. В ее составе имеется два слоя: наружный – пластичный и внутренний – ригидный. Основу клеточной стенки составляет пептидогликан, который ранее называли муреином (лат. mureus – стенка). Он имеется только у эубактерий (кроме микоплазм). Пептидогликан (см. цв. вкл., рис. 4) включает в себя остов и два набора пептидных цепочек – боковых и поперечных. Остов пептидогликана одинаков у всех бактерий и состоит из чередующихся молекул аминосахаров – N-ацетилглюкозамина (N-АцГлю) и N-ацетилмураминовой кислоты (N-АцМур), связанных между собой β-гликозидными связями (рис. 5). Боковые цепочки в каждой молекуле пептидогликана представлены набором идентичных тетрапептидов. Поперечные цепочки также представлены набором из идентичных для данной молекулы пептидогликана пентапептидов, содержащих глицин, – пентаглицинов, однако у разных видов бактерий боковые и поперечные пептиды различны. В тетрапептидной боковой цепочке у большинства грамотрицательных бактерий имеется диаминопимелиновая (диаминопимеловая) кислота (ДАП) – уникальный компонент клеточной стенки, обнаруженный только у прокариот. Кроме того, в составе боковых цепочек пептидогликана обнаружены D-аминокислоты (D-аланин, D-глутамин). Боковые тетрапептиды связаны с N-ацетилмураминовой кислотой остова. Связывание боковых тетрапептидов между собой происходит путем образования поперечных пентаглициновых мостиков между D-аланином одной цепи и диаминопимелиновой кислотой (или иной аминокислотой) другого бокового пептида. Наличие двух типов связей (гликозидные и пептидные), которые соединяют субъединицы пептидогликанов, придает этому гетерополимеру структуру молекулярной сети (см. цв. вкл., рис. 4). Благодаря этим связям пептидогликановый слой клеточной стенки образует огромного размера ригидную мешковидную макромолекулу, которая окружает протопласт, уравновешивает его тургорное давление (у E. coli – до 15 атм.) и придает ему определенную постоянную форму. Пептидогликан может разрушаться под действием различных ферментов, а его синтез блокируют бета-лактамные антибиотики.
Связь между N-ацетилмураминовой кислотой и N-ацетилглюкозамином разрушается лизоцимом, связь между N-ацетилмураминовой кислотой и боковым пептидом (его L-аланином) расщепляют амидазы, а связи межпептидные – эндопептидазы. Пентаглициновый мостик стафилококкового пептидогликана разрушается лизостафином. Образование поперечных сшивок между боковыми цепочками тетрапептидов блокируется пенициллинами (бета-лактамными антибиотиками). Это приводит к разрыхлению пептидогликановой сети, следствием чего является осмотический лизис растущих клеток. Пептидогликан, помимо того что он определяет постоянную форму бактерий, обладает следующими важнейшими иммунобиологическими свойствами.
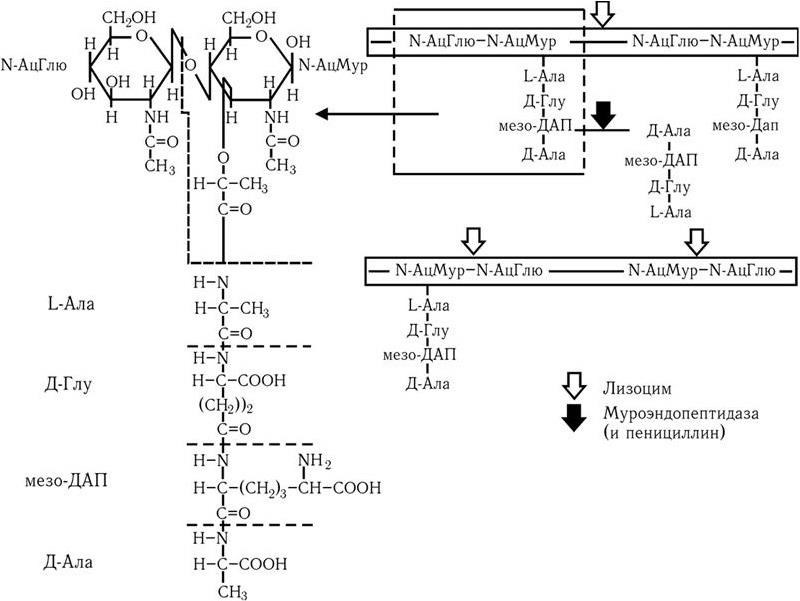
Рис. 5. Химическая структура пептидогликана.
Стрелками указаны участки молекулы, атакуемые лизоцимом, а также муроэндопептидазой и пенициллином. Объяснение в тексте
1. В его составе обнаружены родоспецифические антигенные детерминанты. Они содержатся в гликановом остове и в тетрапептидах. В межпептидных мостиках имеются видоспецифические антигенные детерминанты.
2. Пептидогликан запускает классический и альтернативный пути активации системы комплемента.
3. Он тормозит фагоцитарную активность макрофагов, т. е. защищает бактерии, особенно грамположительные, от фагоцитоза.
4. Угнетает миграцию макрофагов.
5. Способен индуцировать развитие гиперчувствительности замедленного действия.
6. Обладает противоопухолевым действием.
7. Оказывает пирогенное действие на организм человека и животных.
Таким образом, клеточная стенка является чрезвычайно важной биологической структурой бактерий, определяющей многие их специфические свойства. Как отмечалось выше, все бактерии, в зависимости от их отношения к окраске по Граму, делятся на грамположительные и грамотрицательные. Суть окраски по Граму заключается в том, что вначале бактерии окрашивают кристаллическим или генциановым фиолетовым, а затем – раствором Люголя, после чего мазок обрабатывают спиртом и докрашивают водным фуксином. Грамотрицательные бактерии обесцвечиваются спиртом и поэтому окрашиваются в красный цвет, а грамположительные не обесцвечиваются и сохраняют фиолетовую окраску. Это свойство грамположительных бактерий зависит исключительно от особенностей химического состава и структуры их клеточных стенок, так как при разрушении клеточных стенок или утрате их (в случае L-трансформации) они становятся грамотрицательными.
Причину различного отношения бактерий к окраске по Граму объясняют тем, что после обработки раствором Люголя образуется не растворимый в спирте комплекс йода с генциановым фиолетовым, который у грамположительных бактерий в связи со слабой проницаемостью их стенки не может диффундировать из клетки, в то время как у грамотрицательных легко удаляется при промывании их этанолом, а затем водой.
Особенности клеточной стенки грамположительных бактерий
Клеточная стенка грамположительных бактерий имеет однородную структуру, пластичный слой тонкий и ковалентно связан с ригидным слоем. Она значительно толще, чем у грамотрицательных – ее толщина 20 – 60 нм. Основную массу стенки составляет пептидогликан. Он представлен не 1 – 2 слоями, как у грамотрицательных бактерий, а 5 – 6, на его долю приходится до 90 % сухой массы клеточной стенки. Клеточная стенка содержит много тейхоевых кислот (до 50 % сухого веса ее). Тейхоевые кислоты (греч. teichos – стенка) – растворимые в воде линейные полимеры, содержащие остатки глицерина или рибитола, связанные между собой фосфодиэфирными связями. Тейхоевые кислоты – главные поверхностные антигены многих грамположительных бактерий. Они в значительном количестве располагаются между цитоплазматической мембраной и слоем пептидогликана и через поры в нем выступают наружу. Функция тейхоевых кислот полностью не выяснена. Клеточная стенка большинства грамположительных бактерий не содержит липидов, однако у микобактерий и коринебактерий в ней имеются токсические гликолипиды.
Особенность пептидогликанов грамположительных бактерий – частое отсутствие в них диаминопимелиновой кислоты. В клеточной стенке грамположительных бактерий отсутствуют липополисахариды; содержание белка в них сильно варьирует. Белки во многом определяют антигенную специфичность таких бактерий. Например, стрептококки серогруппы А по белкам М и Т подразделяют на несколько десятков серотипов.