Полная версия
Общая и частная гистология
Внутренняя ядерная мембрана содержит сеть переплетающихся промежуточных (виментиновых) филаментов, связанных с ядерной пластинкой, к которой прикрепляются интерфазные хромосомы. Ядерная пластинка состоит из переплетенных промежуточных филаментов (ламинов) толщиной 80—100 нм, образующих кариоскелет.
Ядерные поры – это каналы связи диаметром 70—100 нм между ядром и цитоплазмой, их число и распределение изменчивы. Двусторонний транспорт через пору обеспечивается белками экспортинами (транспортируют РНК из ядра) и импортинами (переносят белковые субъединицы рибосом).
Ядерная пора изнутри выстлана специализированными структурами, образующими комплекс ядерной поры.
Комплекс ядерной поры (КЯП) состоит из немембранных структур: белка-рецептора на сигналы ядерного импорта, а также крупных белковых гранул, определяющих границы поры.
Пора ограничена восемью вертикальными белковыми комплексами, которые представляют собой крупные белковые молекулы или компоненты рибосом, находящиеся в процессе транспорта. По горизонтали пора ограничена тремя кольцевидными структурами, располагающимися одна над другой и соединенными вертикально 8 «спицами». Пора содержит также цитоплазменные волокна, транспортер и ядерную корзинку.
Первое кольцо со стороны цитоплазмы имеет волокнистую структуру – специализированный связывающий белок, который тянется в цитоплазму и обеспечивает импорт в ядро различных субстратов.
Среднее кольцо состоит из восьми трансмембранных белковых молекул, которые выбухают и в просвет поры, и в перинуклеарную цистерну. Эти молекулы прикрепляют гликопротеиновые компоненты КЯП к наружному ободку поры. Центр среднего кольца занят «транспортером», прикрепленным к периферическим белкам среднего кольца.
Третье кольцо со стороны нуклеоплазмы аналогично по строению первому. От него в сторону нуклеоплазмы отходит ядерная корзина, которая деформируется при транспорте веществ через пору.
Ядрышко (nucleolus) – хорошо определяемое только в интерфазе ядерное образование (одно или несколько), наблюдаемое в клетках, активно синтезирующих белок. Под электронным микроскопом в ядрышке выделяют три типичных компонента: фибрилшрный компонент, состоящий из тонких, диаметром 5–8 нм, нитей (совокупность первичных транскриптов – предшественников рРНК); гранулярный компонент – скопление плотных частиц диаметром 10–20 нм (поздние стадии образования предшественников рРНК); аморфный компонент, представляющий собой связанный с ядрышком хроматин, состоящий из ДНК в области ядрышкового организатора хромосомы.
Фибриллярный и гранулярный компоненты ядрышка образуют ядрышковую нить (нуклеолонема) толщиной 60–80 нм, формирующую в пределах ядрышка широкопетлистую сеть.
Ядрышко участвует в синтезе рРНК и формировании предшественников рибосомальных субъединиц. Размеры и число ядрышек увеличиваются при повышении функциональной активности клетки.
Плазматическая мембрана (плазмолемма, клеточная мембрана) окружает клетку и ограничивает ее от внешней среды; обеспечивает распознавание клеткой других клеток, а также взаимодействие с межклеточным веществом (прикрепление к его элементам, взаимодействие с сигнальными молекулами: гормонами, медиаторами, цитокинами и др.); регулирует движение ионов и макромолекул из клетки и в клетку (селективная проницаемость), осуществляет активный и пассивный транспорт веществ (эндоцитоз – фагоцитоз и пиноцитоз, экзоцитоз); обеспечивает механическое и химическое взаимодействие между клетками, а также движение клетки (образование псевдо-, фило– и ламеллоподий).
Молекулярное строение плазматической мембраны описывается жидкостно-мозаичной моделью, согласно которой она состоит из двойного фосфолипидного слоя, внутри которого распределены интегральные и периферические белки; гидрофильные концы фосфолипидов обращены наружу, гидрофобные цепи – внутрь; между хвостами противолежащих молекул фосфолипидов имеются слабые гидрофобные связи.
При замораживании – скалывании плазмолемма расщепляется вдоль так, что большая часть интегральных белков отходит к внутреннему листку (Р-поверхность, или протоплазматическая) и только некоторые – к наружному (Е-поверхность, или наружная).
Белки занимают разное положение в бислое, составляя более 50 % от массы мембраны. Некоторые белки прикреплены к мембране с помощью специальных компонентов цитоскелета (интегральные белки), другие передвигаются к поверхностям мембраны (трансмембранные белки – белки-переносчики, белки мембранных насосов, белки ионных каналов).
В плазмолемме присутствуют также гликолипиды, участвующие в межклеточных взаимодействиях, холестерин, обеспечивающий стабильность ее структуры, а также молекулы углеводов, соединенные либо с гликолипидами, либо с интегральными белками и действующие как рецепторы химического взаимодействия между клетками (например, рецепторы гормонов). В составе плазмолеммы имеются также интегрины – трансмембранные белки, служащие рецепторами для внеклеточных макромолекул (фибронектина и ламинина), которые обеспечивают связь клетки с внеклеточным матриксом.
Гликокаликс (поверхностная оболочка) находится на внешней поверхности плазматической мембраны. Он сформирован олигосахаридными компонентами интегральных гликопротеинов мембраны и гликолипидами, придает отрицательный заряд поверхности клетки, играет роль в иммунологической специфичности, содержит антигены групп крови, участки-рецепторы, а также служит защитным механическим барьером. В клетках тонкой кишки гликокаликс содержит ферменты, гидролизующие дисахариды и полипептиды, участвующие в пищеварении.
Подмембранный (кортикальный) слой плазмолеммы образован упорядоченной сетью поперечно связанных белковых нитей из актина и актинсвязанных белков (прежде всего филамина), которая выстилает изнутри Р-поверхность плазматической мембраны.
Через плазмолемму постоянно осуществляется трансмембранный транспорт, который бывает пассивным и активным.
Пассивный транспорт происходит без затрат энергии, за счет градиента концентрации, и включает простую и облегченную диффузию. Простая диффузия – перенос мелких неполярных молекул (кислород, азот, углекислый газ, бензол) и незаряженных полярных молекул (вода, глицерин) по градиенту концентрации. Облегченная диффузия – прохождение большинства ионов и мелких молекул через мембрану по специальным белковым каналам или с помощью белков-переносчиков.
Активный транспорт является энергоемким процессом, в котором перенос более крупных молекул осуществляется с помощью белков-переносчиков против градиента концентрации ионов, с активным участием плазмолеммы (эндо– и экзоцитоз).
Эндоцитоз включает пиноцитоз – перенос жидкости и мелких молекул диаметром 60—100 нм (неселективный и селективный) и фагоцитоз – поглощение веществ с образованием крупных эндосом (диаметром 250 нм и более).
Неселективный эндоцитоз осуществляется путем инвагинации участка плазмолеммы с последующим слиянием его краев и образованием эндосомы. Слияние мембран происходит с образованием активного фузогенного комплекса (ФК). ФК образуется при взаимодействии специального N-этилмалеимидчувствительного белка слияния (NSF) и связывающего белка (SNAPS).
Селективный эндоцитоз, или рецепторно-опосредованный эндоцитоз, – более эффективный способ эндоцитоза (перенос гормонов, фагоцитоз лейкоцитами бактерий), опосредованный путем связывания мембранных рецепторов с молекулами поглощаемого вещества с образованием окаймленных пузырьков и кавеол.
Участок мембраны, несущий рецепторы с макромолекулами поглощаемого вещества – лигандами, формирует окаймленную ямку, которая окружается гексагональной корзиной из окаймляющего белка клатрина. Клатринная корзина усиливает инвагинацию, превращая ямку в окаймленный пузырек (ОП). Как только ОП «проваливается» в цитоплазму, клатрин сбрасывается с его поверхности и возвращается на поверхность клетки, снова встраиваясь в плазмолемму, а содержимое ОП подвергается процессингу (переваривание).
Экзоцитоз – это выведение продуктов из клетки без нарушения целостности плазмолеммы. Избыток мембраны удаляется при экзоцитозе, что предотвращает бесконечное увеличение поверхности клетки.
Трансцитоз характерен для некоторых типов клеток, например эндотелиоцитов стенок кровеносных сосудов, особенно капилляров. Он объединяет признаки эндоцитоза и экзоцитоза. Трансцитоз заключается в том, что на одной поверхности клетки формируется эндоцитозный пузырек, который переносится к противоположной ее поверхности и, становясь экзоцитозным пузырьком, выделяет свое содержимое во внеклеточное пространство.
Цитоплазма – второй важнейший компонент клетки, в котором располагаются все органеллы (рибосомы, матохондрии, комплекс Гольджи и т. д.) и совершаются физиологические процессы.
Рибосомы (ribosomae) – плотные немембранные органеллы, представляющие собой частицы рибонуклеопротеина (рРНК и ряд белков) размером 12 х 25 нм, состоящие из асимметричных большой и малой субъединиц. Они часто группируются вдоль нити иРНК, формируя полисомы; соединившись с иРНК, синтезируют белок.
Рибосомы и полисомы могут быть свободными (синтезируют белки для клетки) или фиксированными на мембранах ЭПС (синтез белка для выделения из клеток).
Гранулярная ЭПС – система трубочек и цистерн, окруженных мембраной, снаружи усеянной рибосомами. Она обеспечивает биосинтез всех мембранных белков и белков, предназначенных для экспорта из клетки, а также начальное гликозилирование и посттрансляционные изменения белковых молекул.
В ГЭПС различают внутреннюю часть (цистерну), содержащую продукты синтеза, и рецепторы (специфические гликопротеины, рибофорины) на мембранах, к которым прикрепляются большие субъединицы рибосом.
иРНК располагается между малой и большой субъединицами рибосомы; наблюдается в клетках, синтезирующих белки «на экспорт» (гландулоциты слюнных желез, желез пищеварительного тракта и др.).
Агранулярная (гладкая) ЭПС – нерегулярная, неправильной формы сеть анастомозирующих трубочек, канальцев, цистерн и везикул диаметром 20—100 нм, окруженных мембранами без рибосом. Она выполняет функции небелкового синтеза (углеводы, липиды, холестерин), синтеза стероидных гормонов, детоксикации лекарств, обмена жиров и холестерина, выделения и обратного поглощения ионов кальция во время сокращения и расслабления миофибрилл.
Переходная (транзиторная) ЭПС – участок перехода ГЭПС в АЭПС у формирующейся поверхности комплекса Гольджи, в котором цистерны распадаются на отдельные окаймленные транспортные пузырьки, переносящие материал из ГЭПС в комплекс Гольджи.
Митохондрии (mitochondriones) – палочковидные, различимые в световом микроскопе мембранные полуавтономные органеллы длиной 2—10 мкм и шириной 0,2–2,0 мкм (рис. 2.2). Они построены из наружной и внутренней мембран, разделенных межмембранным пространством (матрикс).
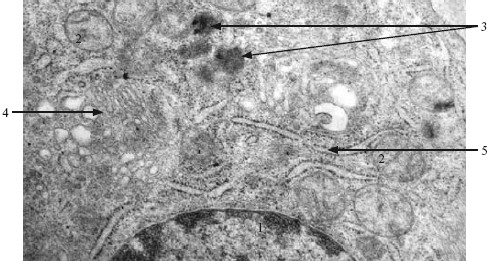
Рис. 2.2. Фрагмент клетки. ТЭМ. ×20 000.
1 – ядро; 2 – митохондрии; 3 – лизосомы; 4 – комплекс Гольджи; 5 – цистерны гранулярной ЭПС.
Наружная мембрана окружает всю органеллу, содержит много молекул специализированных транспортных белков (поринов), а также небольшое количество рецепторов и ферментных систем.
Внутренняя мембрана формирует кристы, содержащие ферментные комплексы цепи переноса электронов, которые участвуют в окислительном фосфорилировании. В ее состав входят белки 3 типов: 1) транспортные, 2) ферменты дыхательной цепи и сукцинатдегидрогеназа, 3) комплекс АТФ-синтетазы.
Кристы – это складки внутренней мембраны толщиной 18–20 нм. На них находятся элементарные (грибовидные) частицы – оксисомы, или F1-частицы, состоящие из головки диаметром 8–9 нм и ножки толщиной 2–3 нм, на которых происходит сопряжение процессов окисления и фосфорилирования.
Большинство крист по форме пластинчатые (ламеллярные), в некоторых клетках (клетки коркового вещества надпочечника, желтого тела яичника и др.) они имеют форму пузырьков и трубочек (тубулярно-везикулярные кристы).
Матрикс (внутренняя среда) содержит гранулы, которые связывают двухвалентные катионы (магния и кальция). В матриксе находятся ферменты цикла Кребса, ферменты белкового синтеза и окисления жирных кислот.
Митохондрии имеют собственный генетический аппарат из ДНК (кольцевой формы), иРНК, тРНК и рРНК (с ограниченной способностью к кодированию), поэтому большинство белков митохондрий кодируются ядерной ДНК. Они производят аденозин-трифосфат (АТФ) – главный и первичный запас энергии в клетке. Это энергетические станции клетки.
Жизненный цикл митохондрий – около 9—10 сут, их разрушение происходит путем аутофагии, а образование новых – путем перешнуровки предсуществующих.
Комплекс Гольджи (КГ) – мембранная органелла, которая состоит из нескольких дисковидных мешочков (цистерн), собранных в стопку, не анастомозирующих между собой, а также пузырьков и вакуолей (рис. 2.3).
Другая сторона стопки образует внешнюю вогнутую поверхность (транс-поверхность, или поверхность созревания).
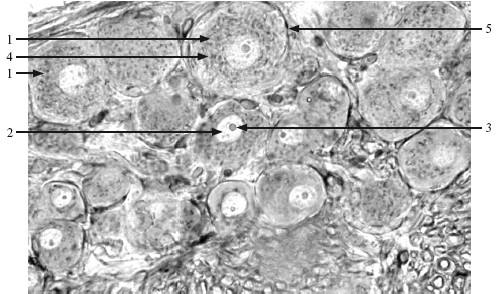
Рис. 2.3. Комплекс Гольджи. ×300.
1 – нейроцит (псевдоуниполярный); 2 – ядро; 3 – ядрышко; 4 – комплекс Гольджи; 5 – олигодендроглиоцит.
Мелкие везикулы диаметром 40–80 нм (включая транспортные везикулы ЭПС) связаны с внутренней выпуклой цис-поверхностью, а конденсированные вакуоли – секретированные вещества, конденсированные в гранулы, – с внешней транс-поверхностью.
В зависимости от типа клетки и ее активности размеры и степень развития КГ варьируют.
Функции КГ: переработка и перераспределение мембран; синтез полисахаридов и гликопротеинов; модификация продуктов ЭПС путем прибавления жирных кислот, сульфатирования, гликозилирования; концентрация и упаковка синтезированных веществ в секреторные гранулы; участие в образовании лизосом.
Лизосомы (lysosomae) – плотные органеллы, окруженные одинарной мембраной (см. рис. 4.2). Они содержат около 60 гидролитических ферментов (протеазы, нуклеазы, липазы, гликозидазы, фосфорилазы, фосфатазы, сульфатазы), активно участвующих во внутриклеточном пищеварении. Лизосомы могут быть обнаружены по позитивной реакции на кислую фосфатазу.
Первичные лизосомы – вновь образованные тельца, еще не принимавшие участия в пищеварении.
Вторичные лизосомы – органеллы, в которых происходит переваривание; они имеют различное происхождение. Различают следующие разновидности лизосом:
– гетерофагическая вакуоль, или фаголизосома, формируется, когда вещества, поглощенные из внешней среды, изолируются в фагосоме, которая сливается с первичной лизосомой;
– аутофагическая вакуоль, или аутофагосома, образуется, когда органелла, подлежащая разрушению, окружается мембранами ГЭПС, которые формируют вакуоль, сливающуюся с первичной лизосомой;
– мультивезикулярное тельце образуется, когда жидкость, поступившая в клетку внутри мелких пиноцитозных пузырьков, окружается мембраной и формируется вакуоль, сливающаяся с первичной лизосомой;
– остаточные тельца – лизосомы, содержащие непереваренные вещества.
Лизосомы формируются гранулярной ЭПС и КГ.
Гидролазы лизосом, возможно, движутся прямо из элементов ЭПС в первичные лизосомы в обход КГ.
Распространенным типом остаточных телец в организме человека являются липофусциновые гранулы, накапливающиеся в некоторых клетках (нейроны, кардиомиоциты) при старении.
Эндосомы, или окаймленные пузырьки, вовлечены в связанный с рецепторами плазмолеммы захват клеткой специфических макромолекул из окружающей среды и их переваривание. Они формируются после того, как специфические макромолекулы связываются с рецепторами плазматической мембраны, что вызывает скопление рецепторов в одном месте и формирование покрытых плазмолеммой углублений, которые инвагинируют и отделяются, образуя окаймленные пузырьки, окруженные клатрином.
Клатрин формирует структуру, похожую на решетчатую корзинку. Последняя окружает везикулу предположительно для того, чтобы эндосомы не сливались с другими мембранными органеллами.
Выделяют ранние (периферические) и поздние (перинуклеарные) эндосомы.
Эндосомы обеспечивают перенос макромолекул с поверхности клетки в лизосомы и их частичный или полный гидролиз на стадиях, предшествующих лизосомальному уровню деградации.
Функция лизосом: активное участие в завершающих этапах процесса внутриклеточного переваривания захваченных клеткой макромолекул, что лежит в основе гетерофагии (защитные реакции клетки) и аутофагии (омоложение, т. е. обновление клеточных структур).
Пероксисомы (микротельца) (peroxysomae) – мембранные органеллы, содержащие каталазу – фермент, синтезирующий и разрушающий перекись водорода, которая обладает сильным повреждающим эффектом.
Это сферические или удлиненные пузырьки диаметром 0,05– 1,5 мкм, с умеренно плотным однородным или мелкозернистым содержимым (матриксом), в котором иногда выявляется плотная сердцевина (нуклеоид), имеющая кристаллическое строение.
Выделяют мелкие пероксисомы (микропероксисомы) диаметром 0,05—0,25 мкм, встречающиеся во всех клетках, и крупные (макропероксисомы) – диаметром 0,3–1,5 мкм – в гепатоцитах, макрофагах и других клетках.
Матрикс пероксисом содержит до 15 ферментов. Наиболее важные из них – это пероксидаза, каталаза, оксидаза, уратоксидаза.
Образование пероксисом происходит в гранулярной ЭПС путем отпочковывания от ее элементов, а их ферменты синтезируются в гранулярной ЭПС.
Функции пероксисом: метаболизм перекиси водорода, холестерина, жиров и расщепление пуриновых и пиримидиновых оснований.
Центриоли (centrioli) – немембранные органеллы, которые участвуют в делении клетки (рис. 2.4). Это пара коротких палочек, расположенных под прямым углом друг к другу (диплосома); они образуют клеточный центр (цитоцентр).
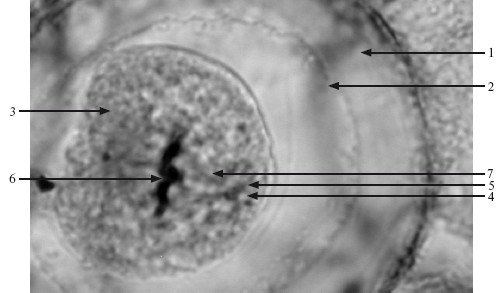
Рис. 2.4. Центросома в яйцеклетке лошадиной аскариды. ×400.
1 – оболочка; 2 – зона сморщивания; 3 – цитоплазма; 4 – центриоль; 5 – центросфера; 6 – хромосомы; 7 – нити ахроматинового веретена.
Перед делением клетки центриоли самоудваиваются: каждая родительская центриоль формирует под прямым углом к себе дочернюю центриоль.
Центриоли образуют полюса митотического веретена, где микротрубочки берут начало и сходятся.
Каждая центриоль имеет стенку, состоящую из 9 триплетов микротрубочек (на поперечном срезе центриоли они напоминают колесо), связанных поперечными белковыми мостиками («ручками»). Каждый триплет центриоли связан со сферическими тельцами диаметром 70–75 нм (сателлитами); расходящиеся от них микротрубочки образуют центросферу.
Формируют базальные тельца, от которых отходят реснички и жгутики – органеллы специального значения, участвующие в процессах движения. Их основу составляет каркас из микротрубочек, называемый осевой нитью, или аксонемой.
Аксонема образована девятью периферическими парами микротрубочек и одной центрально расположенной парой, окруженной центральной оболочкой, от которой к периферическим дублетам расходятся радиальные спицы. Периферические дуплеты связаны друг с другом мостиками нексина, а к соседним дуплетам отдают «ручки» из белка динеина, который обладает активностью АТФазы.
Цитоскелет (cytoskeleton) относится к структурному каркасу клетки. Это компоненты цитоплазмы, которые поддерживают форму клетки, стабилизируют прикрепление клетки, лежат в основе эндо– и экзоцитоза, играют роль в подвижности клетки и т. д.
В цитоскелет входит несколько волокнистых структур: микротрубочки, микрофиламенты, промежуточные филаменты и микротрабекулы.
Микротрубочки – прямые структуры диаметром 25 нм и длиной несколько микрометров; толщина стенки составляет 4–5 нм, а просвет 14–15 нм. Различают 2 вида микротрубочек:
– лабильная популяция находится в цитоплазме в свободном состоянии и полимеризуется или деполимеризуется в зависимости от температуры, давления, наличия каких-либо лекарств и т. д.;
– стабильная популяция формирует стенки центриолей и аксонемы ресничек и жгутиков; имеет стенку толщиной 4–5 нм, которая окружает внутреннюю полость и состоит из 13 параллельных спирально расположенных протофиламентов (линейных полимеров тубулина).
Микротрубочки часто заканчиваются около центриолей в маленьких плотных тельцах (сателлиты центриолей).
Функции микротрубочек: поддержание формы и полярности клетки и внутриклеточного транспорта макромолекул в ней, обеспечение движения ресничек, жгутиков и хромосом (в митозе).
Актиновые микрофиламенты (тонкие филаменты) – филаменты толщиной 5–6 нм (F-актиновая форма), которые содержат 10–15 % от общего количества белка в клетке; актин существует также в глобулярной форме (G-актин). Они многочисленны на периферии клетки, где формируют под плазматической мембраной плотную сеть. Участвуют в трансформации цитоплазмы в формы зольгель, эндоцитозе, экзоцитозе, а также локомоции немышечных клеток.
Миозиновые филаменты (толстые филаменты) диаметром в среднем 14–15 нм. Обычно ассоциированы с актином в мышечных клетках. В поперечнополосатых мышцах полимеризуются в ясно различимые филаменты.
Миозин также находится в низких концентрациях в немышечных клетках, однако его функциональная роль здесь не совсем ясна.
Промежуточные филаменты – это гетерогенная популяция, включающая филаменты диаметром от 8 до 11 нм.
Выделяют кератиновые, виментиновые, десминовые, нейро-и глиальные филаменты.
Кератиновые филаменты (тонофиламенты) обычно располагаются в эпителиальных клетках и часто ассоциированы с десмосомами.
Десминовые филаменты формируют в скелетных, гладких и сердечной мышцах сети, которые объединяют миофибриллы.
Виментиновые филаменты присутствуют в фибробластах и других клетках – производных мезенхимы. Они стабилизируют содержимое ядра и тесно ассоциированы с ядерной оболочкой и ядерными порами.
Нейрофиламенты осуществляют поддержку отростков нейронов и обеспечивают состояние геля в цитоплазме клеток.
Глиальные филаменты присутствуют в астроцитах, олигодендроцитах и клетках микроглии центральной нервной системы (ЦНС).
Микротрабекулярная решетка (МР) – трехмерная сеть нитей в эргастоплазме некоторых клеток, обнаруживается только под электронным микроскопом.
Наличие этой решетки указывает на то, что эргастоплазма – не просто гомогенный белковый раствор, но является в высшей степени структурированным гелем, который объединяет филаментные компоненты и органеллы в единое структурно-функциональное целое.
Предполагают, что МР участвует в координации метаболической активности компонентов клетки с помощью специальных «управляющих» протеинов.
Включения (inclusiones) – скопления некоторых временно присутствующих веществ внутри клетки. Обычно к ним относятся скопление гликогена, капли липидов и секреторные гранулы.
Гликоген образует скопления в виде электронно-плотных агрегатов, известных как α-розетки, или в виде мелких кластеров β-частиц.
Жировые капли в зависимости от способа фиксации видны в виде черных (осмий) или светло-серых (альдегиды) образований. Они могут иметь ограничивающую мембрану, но чаще встречаются в виде гомогенной субстанции.
Секреторные гранулы включают слизистые капли, некоторые гормоны, протеины и пигментные гранулы.
2.1. Клеточный цикл
Клеточный цикл (КЦ; cyclus cellularis) – совокупность явлений между двумя последовательными делениями клетки или между ее образованием и гибелью (рис. 2.5).
В ходе КЦ обеспечивается функция воспроизведения клеток и передачи генетической информации. КЦ включает собственно митотическое деление и интерфазу – промежуток между делениями.
Интерфаза включает пресинтетический, или постмитотический (G1), синтетический (S) и постсинтетический, или премитотический (G2), периоды. В интерфазе клетка увеличивается в размерах и удваивает генетический материал.
В большинстве тканей делится лишь небольшая часть клеток, остальные дифференцируются и пребывают в G0-периоде.
G1-период – промежуток сразу после митотического деления клетки; характеризуется активным ростом клетки и синтезом белка и РНК, благодаря чему дочерние клетки достигают нормальных размеров и восстанавливают необходимый набор органелл. В этот период синтезируются особые «запускающие белки», или активаторы S-периода, которые обеспечивают переход клетки в S-период.