


Полная версия
Заметки о древних и современных этносах
В первой половине IX в. до н.э. часть киммерийского населения из-за ухудшения природно-климатических условий мигрировала из степей Северного Причерноморья в Карпато-Балканский регион, на Большую Венгерскую равнину, где в результате с частью местного населения, в первую очередь с женщинами, сформировался местный вариант киммерийской культуры, известный под названием культуры Мезёчат. Отсюда киммерийцы стали проникать в другие регионы Центральной и Западной Европы. «Кочевой или полукочевой образ жизни мезёчатского населения помогал распространению восточной идеологии и технических достижений киммерийцев (конский убор, предметы вооружения и инсигнии власти) далее на запад. Заимствование киммерийского вооружения и узды происходило как в результате мирных контактов с западной группой киммерийцев, так и в результате их военных набегов. О киммерийских грабительских набегах свидетельствуют поселения со следами разрушений и находками наконечников стрел киммерийского типа из Моравии и Нижней Австрии, а также, возможно, укрепленные поселки в западной Венгрии, юго-восточной Австрии и в Задунавье» [136, с.382—383].
В середине VIII в. до н.э. следующий пик аридизации климата вместе с длительными засухами и вызванными ими экономическим кризисом, вынудил киммерийцев совершать грабительские походы в Закавказье и страны Древнего Востока. «Именно с момента начала грабительских походов во второй половине VIII в. до н.э. о киммерийцах начинают сообщать письменные источники: ассиро-вавилонские документы и греческие авторы» [136, с.383].
В начале VII в. до н.э. когда значительная часть киммерийских воинов находилась в Передней Азии, в восточноевропейские степи с востока приходит первая волна нового кочевого населения, известного под именем скифов. С.В.Махортых (2008) предполагает, что причиной продвижения скифов на запад, была аридизация второй половины VIII в. до н.э., вынудившая их искать новые пастбища. Наиболее благоприятные условия на своем пути они встретили в Предкавказье, а затем стали проникать в украинскую лесостепь. После ряда столкновений с местным киммерийским и кобанским населением, скифы, по-видимому, затем установили с ними мирные отношения.
В 70-е годы VII в. до н.э. в степях Восточной Европы появилась вторая волна скифов, которая вытеснила киммерийцев с Северного Кавказа, которые частично ушли в степные и лесостепные районы Украины, частично – за Дунай на территорию фракийцев—треров. Столкновения со скифами в Малой Азии и в степях Причерноморья заставили киммерийцев отступить в Каппадокию. Там, а также у г. Синоп на южном берегу Черного моря они обитали до конца VII в. до н.э., когда окончательно были разбиты лидийским царем Алиаттом [136].
Язык киммерийцев неизвестен. От него сохранились имена трёх царей в ассирийской транскрипции (Andak atru, Dugdamme, Teu pa).
В настоящее время, как считают генетики, им удалось получить палеоДНК киммерийцев (двое мужчин и женщина).
Гаплогруппы мужчин: R1b1a + H9a, Q1a1 + C5c, женщина – R [251].
Судя по гаплогруппам, киммерийцы пришли с востока, т.к. мтДНК С5с и Y-хромосомная Q1a1 родом из Азии. В частности, древняя Q1a1 возрастом 3 тыс. лет обнаружена в Китае [290].
Гаплогруппа Q1-F903 определена у образца AG-2 (16,7 тыс. лет до настоящего времени) со стоянки Афонтова гора в Красноярске [281].
В исследовании Krzewinska et al. (2018) показано, что киммериец с гаплогруппой R1b1a по аутосомно-геномным данным близок к поздним сарматам из Чёрного Яра (Астраханская обл) и Темясово (Башкортостан), некоторым скифам и одному представителю срубно-алакульской культуры. Остальные киммерийцы близки представителям культур железного века Алтайского края, алды-бельской культуры Тувы, Зевакино-Чиликта Восточного Казахстана и Западной Монголии. Киммерийцы сближаются с населением карасукской культуры Минусинской впадины, а не с территориально близкими им популяциями срубной и алакульской культур [251].
Таким образом, можно полагать, что киммерийцы пришли в Причерноморье из Южной Сибири.
1.15. Китойская археологическая культура
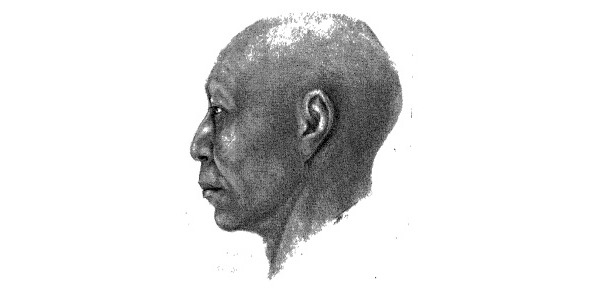
Реконструкция по черепу мужчины (I, 17, 4629) из сада «Локомотив» («Циклодром»). Китойская культура. М. М. Герасимов [45]
В поисках предков по женской линии добрался до берегов озера Байкал, а именно до китойской культуры – моя мтДНК гаплогруппа G2a1 (у литовских татар мтДНК G2a1 распространена с частотой 13,8% и она филогенетически наиболее близка обнаруженной у одного тувинца (Южная Сибирь) и двух якутов [148]) обнаружена у людей из ранненеолитического могильника Шаманка II и, возможно, Локомотив.
Шаманка II расположена на юго-западном побережье оз. Байкал, на мысу Шаманский, в 2 км к юго-востоку от пос. Култук (Слюдянский р-н Иркутской области). Археологически здесь изучено более 110 погребений раннего неолита и раннего бронзового века. Ранненеолитические погребения составляют подавляющее большинство – 92 комплекса, в которых выявлены останки 156 индивидуумов. По данным радиоуглеродного анализа они датируются V тыс. до н.э. [139].
В исследовании Moussa et al. (2015) [260] в Шаманке у 27 индивидов определили следующие мтДНК гаплогруппы: А 18,5%, С 18,5%, D 40,7%, F 11,1%, G2a 7,4%, U5a (5500—4885 BC) 3,7%. У девяти мужчин с Шаманки были определены Y-хромосомные и мтДНК гаплогруппы: K* – D, K* – D, K* – D, K – D, K* – A, K* – G2a, K – C, K —C, K – C. В могильнике Локомотив определены мтДНК гаплогруппы A 13,3%, C 3,3%, D 20%, F 50%, G2a 3,3%, U5a 6,7, другие 3,3%. У 7-ми мужчин определены Y-ДНК гаплогруппы: R1a1-М217 (5500—4885 BC) 28,6% (у обоих мтДНК F), K* 28,6% (мтДНК D и др.), K 28,5% (мтДНК C и G2a), C3-М217 (мтДНК А).
К* и К, как видно из другого исследования, проведенного позже (de Barros Damgaard et al. (2018)) – это разные линии Y хромосомной гаплогруппы N.
В исследовании de Barros Damgaard et al. (2018) [233] получены следующие результаты: Y-хромосомные гаплогруппы – Локомотив: C2b1a1 (C-F4015) (6950±60 BP, мтДНК А), N1c (N-M2087.1) (6680 ±60 BP, мтДНК D4+195); Шаманка: NO1 (NO-M214), N (N-M231), N1 (N-M2291), N1c2 (N-L666), N1c2 (N-L666), N1c2 (N-L666); МтДНК гаплогруппы – Локомотив А, D4, D4, D4j; Шаманка C4 (6373 ± 32 BP, Y-ДНК N1-M2291), C4 (6483 ± 37 BP, Y-ДНК NO1-M214), C4, D4a1, D4e1 (6329±33 BP), D4j (7005±40 BP), F1b1 (6386±34 BP), G2a1 (6319±33 BP, Y-ДНК N1c2-L666), G2a1 (6815±38 BP, Y-ДНК N1c2-L666), G2a1 (6886±43 BP).
В настоящее время, по данным de Barros Damgaard et al. (2018) наиболее близкая китойской Y-ДНК гаплогруппа N1c2 встречается у коми и хантов [233] (в целом это гаплогруппа самодийских народов, бывшая N1b).
А.П.Окладников писал, «что в китойское время обнаруживаются признаки несравненно более широких, чем это было в исаковское или серовское время, контактов населения Прибайкалья с другими, нередко весьма отдаленными, территориями как на западе, так и на востоке». Он отмечает, что имеется обширный «материал, свидетельствующий об этих контактах китойской культуры с такими, например, далекими культурами, как ямная и катакомбная, китойских памятников с памятниками типа Мариупольского могильника или могильника Вэстербьерс на острове Готланде – на западе, с культурой яншао в Северном Китае…» [147]
Нужно отметить, что ямная и катакомбная культуры намного младше китойской, но поскольку А.П.Окладников отмечает черты сходства у них, можно полагать, что в состав населения этих культур вошли потомки китойцев, сохранившие некоторые культурные особенности населения Прибайкалья раннего неолита.
Современные генетические исследования, в общем-то, подтверждают это предположение.
Анализ ископаемых ДНК, древностью около 4000 лет указывает на присутствие западно-евразийских линий H, U2e, U5, U7, K (наряду с восточно-евразийскими линиями C4, C5, D, G2a, M5) в северо-западной провинции Китая, на рубеже раннего/среднего неолита [180, с.28—29]
Б.А.Малярчук и др. сообщают о наличии мтДНК гаплогруппы G у европейских народов – коми (A, C, Z, D, G) 11,8%, латыши G 0,3%, русские южных и центральных регионов (M*, C, D, G) 1,3%, русские северо-западного региона (A, Z, D, M*, G) 4%, поляки, чехи (A, N9a, M*, C, D, G) 1,6% [133, с.404].
Н.В.Трофимова и др. сообщают, что гаплогруппа G2a с высокой частотой встречается в популяциях бурят (11%), монголов (8,5%), а в целом гаплогруппа G присутствует также в Волго-Уральском регионе – у коми (2,4%), татар (1,6%), чувашей (0,7%) и башкир (4,8%) [190, с.1468].
Международная группа генетиков обнаружила мтДНК гаплогруппу G2a у поляков-кашубов (Кашубия находится в польском Поморье) (0,5%) и у украинцев (0,6%). Кроме того, они определили другие восточноевразийские гаплогруппы у поляков Подгалья (А – 1%, D – 1%), у украинцев (А – 0,6%, С – 1,2%). При этом, по их мнению, гаплогруппы G2a и С4а1 (18—25 тыс. лет назад) попали в земли, ныне населенные славянами, в верхнем палеолите, а С5 – в неолите [259].
С верхним палеолитом, на мой взгляд, перебор, поскольку в неолитических культурах линейно-ленточной керамики (КЛЛК) и Старчево восточноевразийские мтДНК гаплогруппы A, C, D, G не обнаружены. КЛЛК была распространена на территории нынешней Польши и заходила в Беларусь и Украину.
Гаплогруппа G2a была обнаружена у туймазинских татар (4,2%) и отсутствует у казанских [190, с.1467].
МтДНК гаплогруппа G2 есть у тубаларов 2,1%, манси 6,1%, тувинцев 6,3%, ульчей 1,2% [180, с.15].
Палеогенетики нашли мтДНК гаплогруппу С в могильнике Ясиноватка (около 6440—6080 гг. до н.э.) днепро-донецкой культуры (в двух из шести образцов). В двух образцах определена мтДНК гаплогруппа Т, в одном U3 и в одном, вероятно, U1. Для большинства образцов определен возраст (calBC): C – 5432—5148, С – (не датирован), Т – 5616—5482, Т- не датирован, U3 – 5434—5221, возможно U1 – 5437—5090. Авторы статьи предполагают, что появление гаплогруппы С на территории Украины связано с депопуляцией в Прибайкалье [255]. Гаплогруппы C и C4a2 обнаружены у представителей днепро-донецкой культуры и в другом исследовании [268].
Нужно отметить, что Н.С.Котова относит могильник Ясиноватка 1, наряду с могильником Вовниги 3 и второй и третьей группами Вилнянского могильника к сурской культуре и датирует их 5100—5000 лет до н. э. Она отмечает в их керамике и погребальном обряде влияние традиций второго периода Азово-Днепровской культуры. По мнению Н.С.Котовой, после 5000 лет до н.э. сурское население было ассимилировано носителями поздней Азово-Днепровской и ранней Среднестоговской культур [250].
В ямной культуре из пяти исследованных образцов три относились к мтДНК гаплогруппе U (U, U5, U5a) и два С4 (С4а2 EBA 3990±70 (Ki-11177) /2815—2291; C4a2 EBA 3740±70 (Ki-11201) /2433—1945). По мнению генетиков, гаплогруппа С4а2 в ямной культуре может быть наследием днепро-донецкой культуры. Она сходна с образцом из Мариупольского могильника в Ясиноватке, с образцами современного населения Прибайкалья и шерпы с Тибета [267].
Палеогенетики нашли мтДНК гаплогруппу G2a1 в Заилийском Алатау (Кызылбулак, возраст 3516 (1618—1513 calBCE) вместе с Y-ДНК Q1a2b2 (памятники кульсайской группы Юго-Восточного Семиречья – региональный вариант андроновской (федоровской) культуры)) [264] и в Паннонии у венгра эпохи «завоевания родины».
Китойская культура является наиболее древней ранненеолитической в Прибайкалье, и датируется VII – VI тыс. до н.э. (даты не были калиброваны). Разрыв между китойскими и серовско-исаковскими погребениями составил 700 лет.
Археологи выделяют на побережье озере Байкал в раннем неолите два хронологических периода: ранний (8,2—7,5 тыс. кал. л.н.) и поздний (7,4—7,0 тыс. кал. л.н.) [48, с.139], в Западном Забайкалье выделены следующие хронологические периоды неолита: начальный неолит – 12—7,5 тыс. л. н.; ранний неолит – 7,5—6,0 тыс. л. н.; развитый неолит – 6,0—4,5 тыс. л. н.; поздний неолит – 4,5—3,8 тыс. л. н. и отмечают присутствие двух керамических традиций: сетчатую (с оттисками сетки-плетенки) и шнуровую хайтинского типа [49; 210].
Начальный неолит Западного Забайкалья обнаруживает сходство каменной индустрии с позднепалеолитическими комплексами [210].
Керамику с оттисками сетки-плетенки, характерную для всех объектов периодов раннего неолита (и далее вплоть до позднего неолита), Горюнова и Новиков (2017) рассматривают как автохтонную. На стоянках с многокомпонентными керамическими традициями в комплексах раннего периода преобладают сосуды хайтинского типа, в то время как на поздних этапах раннего неолита они количественно уступают сетчатым и значительно видоизменяются. В связи с этим, археологи предполагают, что в Прибайкалье был приток иной группы населения со своими сложившимися гончарными традициями и поэтому керамику хайтинского типа рассматривают, как привнесенную. Принадлежность этих традиций к разным группам населения подтверждают находки сетчатых и хайтинских сосудов в ранненеолитических захоронениях разных погребальных практик (сетчатая – в китойских, хайтинская – в уюгинском) [48, с.139].
Н.В.Цыденова (2021) отмечает, что «широкое распространение и длительное существование шнуровой и штриховой керамических традиций, погребального обряда близкого китойскому, должны рассматриваться, как свидетельство существования единых традиций в раннем – позднем неолите Западного Забайкалья» [210, с.22].
Традиции, свойственные неолитическому населению Прибайкалья, встречаются и у современного населения этого региона. Так, например, в Локомотиве, в погребении обнаружены останки волка в сочетании с человеческим черепом [14; 154; 230]. До недавнего времени такой обычай встречался у кетов: «Кеты хоронили умерших в земле. Раньше во время похорон у могилы разжигали костер, который тушили, когда могилу закапывали. С покойником укладывали и его вещи: нарты, челнок, нож, трубку (все в сломанном виде); иногда убивали собак. Детей хоронили внутри высокого, расколотого вдоль пня кедра, специально срубленного для этой цели» [95, с.697].
Исследования антропологов показали, что ранненеолитические (китойские) популяции из могильников Шаманка II и Локомотив обладали выраженной генетической спецификой, отличаясь, как друг от друга, так и от прибайкальского населения более поздних эпох. Это могло быть связано как с депопуляцией, произошедшей в ранненеолитическое время, так и с массовым переселением китойского населения Прибайкалья в другие регионы. Отсутствие генетической связи между населением раннего и позднего этапов неолита Прибайкалья может указывать на пришлое, миграционное происхождение популяций позднего этапа.
При этом, люди из Локомотива в Приангарье, проявляют некоторую близость к населению более поздних этапов этого региона, а люди из Шаманки II резко отличается от групп позднего неолита и бронзы, несколько приближаясь лишь к китойской серии Приангарья. Это, по-видимому, является свидетельством того, что на территории Южного Байкала в ранненеолитическое время проживало население, генетически отличное от населения Приангарья и верховьев Лены. В то же время, относительная близость ранненеолитического населения двух регионов Прибайкалья может свидетельствовать о генетических контактах между ними [139, с.61].
Поэтому, можно предположить, что, несмотря на большой временной разрыв между китойским и серовским этапами, китойская группа Приангарья (Локомотив), при всем своем своеобразии, все же внесла некоторый вклад в генофонд населения последующих эпох этого региона. Поскольку люди из Шаманки II, существенно отличаются от более позднего населения Прибайкалья, то это быть свидетельством, как депопуляции, произошедшей в ранненеолитическое время, так и массового переселения, исхода этой группы китойского населения Южного Байкала в другие регионы.
Генетические данные, приведенные выше, свидетельствуют о том, что ранненеолитическое население из Шаманки II ушло на запад и его следы по мтДНК прослеживаются вплоть до Польши и, возможно, Скандинавии.
Антропологи отмечают близость древнеэскимосских серий из могильников Уэлен и Эквен к популяциям глазковского этапа Прибайкалья. Поскольку население, использовавшее некрополь Локомотив, показывает связи с населением глазковской культуры, то вполне вероятно, что эта группа китойцев, через глазковцев, внесла определенный вклад в формирование генофонда древнеэскимосского населения Чукотки [139, с.60]. В связи с этим, вполне вероятно, что население глазковской культуры было эскимосоязычным (ранее (да и сейчас) в научной литературе было распространено мнение, что население глазковской культуры – это языковые предки эвенков и тунгусов в целом.
О присутствии потомков китойцев среди глазковского населения свидетельствует наличие в погребении раннебронзовой эпохи (глазковская культура) в могильнике Шаманка II мтДНК гаплогруппы G2a1. Все мужчины этого периода в данном некрополе имели Y-ДНК гаплогруппу Q1a2a (5 человек) и Q1a2a1с (1 человек). Y-ДНК гаплогруппа Q1a2 обнаружена также в соседней Усть-Иде (3 человека), Q1a2a1с характерна для населения окунёвской культуры Минусинской впадины эпохи бронзы и современных енисейцев (кеты).
Выше показано, что у двух мужчин из семи из могильника Локомотив определена Y-ДНК гаплогруппа R1a1-М217 (5500—4885 BC (тыс. лет до н.э.)) 28,6% (у обоих мтДНК F), т.е. эти люди жили 7500—6885 лет назад. Интересно, в связи с этим, что самая древняя в Европе R1a обнаружена у мезолитического обитателя Южного Оленьего острова, жившего 7500—7000 лет назад (Y-хромосомная гаплогруппа R1a1 (SRY10831.2)). У него также определена мтДНК гаплогруппа C1g [241]. У трех обитателей этого острова определена мтДНК C1f, для которой не найдено соответствий в текущей базе данных ископаемых и современных митохондриальных геномов [234].
Возможно население Южного Оленьего острова участвовало в этногенезе саамов. Поэтому, участием носителей Y-гаплогруппы R1a в формировании населения Южного Оленьего острова, а затем и саамов можно объяснить проблему, некогда поставившую Е.А.Хелимского в тупик. Он писал: «остается неясным – и трудно укладываемым в какую-либо топохронологическую схему – происхождение немногочисленных, но чрезвычайно ярких лексических параллелей между тюркскими, с одной стороны, и прибалтийско-финскими и саамским, с другой стороны, языками (при отсутствии соответствий в других уральских языках), ср. фин. arpa «орудие колдовства»: тюрк. arpa- «колдовать» (ностратическое происхождение сомнительно), приб.-фин. *käkrä «косой»: тюрк. *käkrü «то же» и др. [199].
Нужно отметить, что по Е. Лагеркранцу саамы имеют наиболее близкое языковое родство с хантами, манси и ненцами [252, p. 336], народами у которых преобладает Y-ДНК N1b.
Y-ДНК гаплогруппа R1a обнаружена в культурах боевых топоров, которые датируются периодом 3200 г. до н. э./2300 до н. э. – 2300 г. до н. э./1800 г. до н. э. Наиболее древняя из них имеет возраст около 4600 лет [241] и очень молода по сравнению с обнаруженными в Азии.
Аутосомный портрет моей мамы Байтасовой Розы Ибрагимовны, из Радлинских. В совпаденцах мамы Мария Мейшутович, Станислав Думин и другие люди с татарскими фамилиями, которых я не знаю.
Roza Radlinskaya
Europe 58%, в т. ч. Eastern Europe: West Slavic 41%, Magyar 15%, Finnis – Finland – менее 2%;
Asia 30%, в т. ч. Central Asia – Mongolia 29%, Indian Subcontinent – Eastern India – менее 1%;
Central Siberia – Yakut – менее 1%;
Western Siberia – Western Siberian Plains – менее 1%;
Middle East & North Africa 12% – Middle East: Anatolia, Armenia & Mesopotamia – 12%.
В целом, видимо, правильное отражение картины. У мамы должна быть мтДНК G2a1. Эта гаплогруппа обнаружена, как показано выше, у населения неолитической китойской культуры на оз. Байкал (7 тыс. лет назад) вместе с Y-ДНК NO и N1b. Это означает, что предки мамы были уралоязычными (скорее всего, прасамодийцы).
Ближе к рубежу нашей эры митохондриальная гаплогруппа G2a1 обнаружена в некрополе знатных хунну в Монголии. В одном из исследований у ранних хунну (7 мужчин) G2a1 (G2a1e – два образца) обнаружена вместе с Y-хромосомными гаплогруппами R1b (R-M343), R1 (RP236; R-M173). У поздних хунну (один мужчина из 25) – (G2a+152) вместе с Y-хромосомной гаплогруппой Q1a2a [245]. В другом исследовании (52 древних генома хунну (26 мужчин и 26 женщин) из некрополя Tamir Ulaan Khoshuu (TUK) в Центральной Монголии, I век до н.э. – I век н.э.), определили, что 16 мужчин (некоторые из них родственники) обладали Y-хромосомной гаплогруппой R1a, 5 – Q1a, 2 – N и 1 – G1. Анализ мтДНК выявил 28 митохондриальных гаплотипов, принадлежавших к 11 гаплогруппам, как западноевразийским (H, J, T, U, X), так и восточноевразийским (A, B, C, D, F, G). Все R1a1a гаплотипы принадлежат субкладу R1a1a1b2, азиатской ветви гаплогруппы R1a1a, возникшей в евразийских степях. Большая часть гаплотипов родственна гаплотипам населения Южной Сибири средней бронзы и из региона Красноярска и Тувы железного века, т.е. гаплотипам сибирских скифов и носителей андроновской культуры. В то же время, некоторые гаплотипы хунну (принадлежащие к гаплогруппам Q1a и N1a) родственны гаплотипам аваров и древних мадьяр. Большинство митохондриальных гаплотипов хунну родственны гаплотипам, распространённым к западу от Урала и чаще всего встречались у населения культуры колоколовидных кубков Европы. Некоторые митохондриальные гаплотипы родственны гаплотипам аваров и древних венгров (Keyser et al. 2020) [248].
Митохондриальная гаплогруппа G2a1 обнаружена у древнего венгра периода «взятия родины» в Паннонии (вместе с Y-ДНК гаплогруппой R1b1a1b-M269 (xM412,xU106)). Всего из этого захоронения исследованы 4 человека. У одного из них обнаружена Y-ДНК Q1a-F1096 (xM25) и мтДНК H6a1b, у двух финно-пермская Y-ДНК N1a1a1a1a2-Z1936 (xL1034) и мтДНК N1a1a1a1a. У двух последних определен цвет глаз – голубой, волосы темно- и светлорусые. У носителя Q1a карие глаза и рыжеватые волосы, у R1b – карие глаза и чёрные волосы [265].
Моя бабушка и прабабушка из Шабановичей. У протестированного Шабановича Y-ДНК J, – это переднеазиатская гаплогруппа. Из Радлинских гаплогруппу никто не определял, но их предок был приписан к юшинской хоругви, т.е. они потомки древних усуней, которые были зеленоглазыми и рыжеволосыми, как мой дед Радлинский. Гаплогруппа усуней неизвестна, но скорее всего, это была одна из ветвей R1a.
1.16. Кумсай

Фото антропологической реконструкции (скульптурного бюста) головы по черепу мужчины-вождя ямной культуры. Могильник Кумсай (Актюбинская область, Казахстан). Автор А. И. Нечвалода [128, с.22].
Мужчина зрелого возраста (более 50 лет) был похоронен в сидячем положении, густо осыпан красной охрой (курган 1, погребение 2).
Надпись на фото: «Окончательный вариант реконструкции головы по черепу мужчины ямной культуры. Обращают на себя внимание те черты морфологии черепа, которые нашли отражение в восстановленной голове древнего жителя казахских степей. Это массивное надбровье, глубоко посаженные глаза, мощная нижняя челюсть с тяжелым подбородком»

Портрет, выполненный по антропологической реконструкции (скульптурного бюста) головы по черепу мужчины-вождя ямной культуры. Могильник Кумсай (Актюбинская область, Казахстан). Автор А. И. Нечвалода [128, с.22; 270]
Г. Ф. Дебец связывал краниологический тип черепов ямной культуры из курганов Нижнего Поволжья с морфологическими особенностями черепов верхнего палеолита из Брюнн-Пшедмост. Этот антропологический комплекс был назван «протоевропейский или кроманьонский тип в широком смысле слова» [64].
Большинство представителей ямной культуры из Кумсая имеют рост выше 180 см, а некоторые более 2 м.
Курганный могильник Кумсай в местности Кырык Оба в долине реки Уил, крупнейший в Евразии памятник ямной культуры, насчитывает более 160 курганов. По природной зональности он является самым южным памятником ямного времени. Радиоуглеродные даты Кумсая (Beta №290781 4290±40 BP; ИГАН №4699 3920±70 BP), выполненные по костным образцам человека, представляют интервал в рамках конца IV – первой половины III тыс. до н.э. [202, с.510].
По мнению C.B. Богданова, в материалах Кумсая имеются отчетливые проявления позднеэнеолитической константиновской группы Подонья. В связи с этим Кумсай может представлять собой относительно поздний дериват константиновских памятников, бытовавших в сухих степях Арало-Каспийского района Волго-Уральского варианта ямной культуры параллельно с тамар-уткульскими памятниками, локализованными несколько севернее, в подзоне типичных степей Урало-Каспийского региона [23].



